1.3 LA GÓNADA INDIFERENCIADA
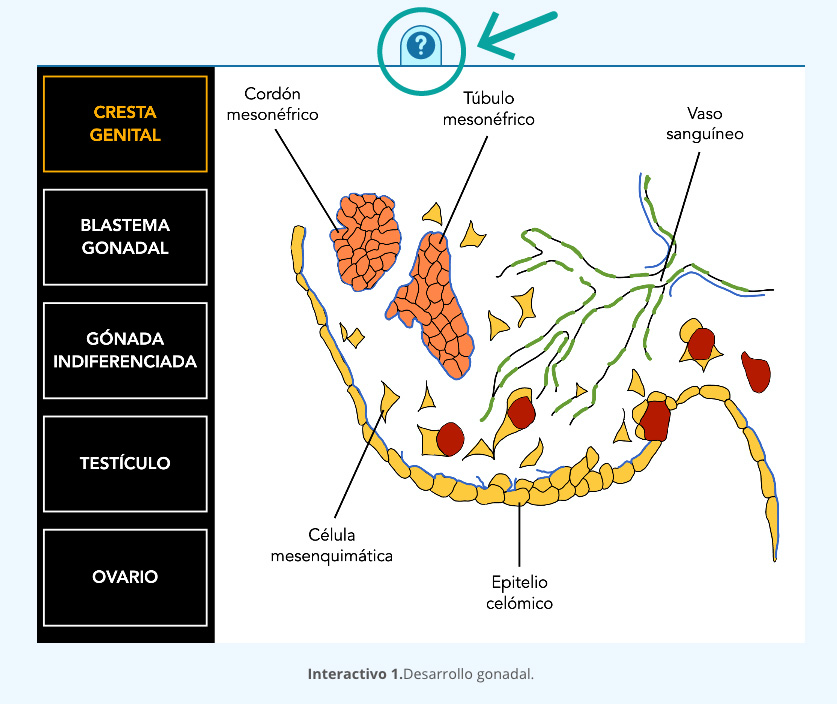
La formación de las crestas genitales en el embrión, inicia como un par de engrosamientos del mesotelio que crecen en la región ventro-medial del mesonefros (riñón fetal). La expresión de los genes Emx2, Wt1, Lhx9 y Sf1 en las crestas genitales es clave para regular los primeros eventos celulares de la morfogénesis gonadal.
En la cresta gonadal, las células somáticas derivadas del mesodermo son de dos tipos: mesenquimáticas y mesoteliales. Ambas inician gran actividad proliferativa junto con las células germinales primordiales (CGPs), dando origen a un proceso de condensación celular en la región ventral del mesonefros denominado “blastema gonadal”.
A partir de este primordio embrionario, se hacen más evidentes los dos tejidos gonadales. En una segunda etapa se establecen los cordones sexuales primarios y el estroma. Los cordones sexuales son arreglos epiteliales que se encuentran delimitados por una membrana basal. Hasta esta etapa las gónadas son estructuralmente indiferenciadas y bipotenciales (interactivo 1).
La gónada primitiva está integrada por dos tipos de tejidos:
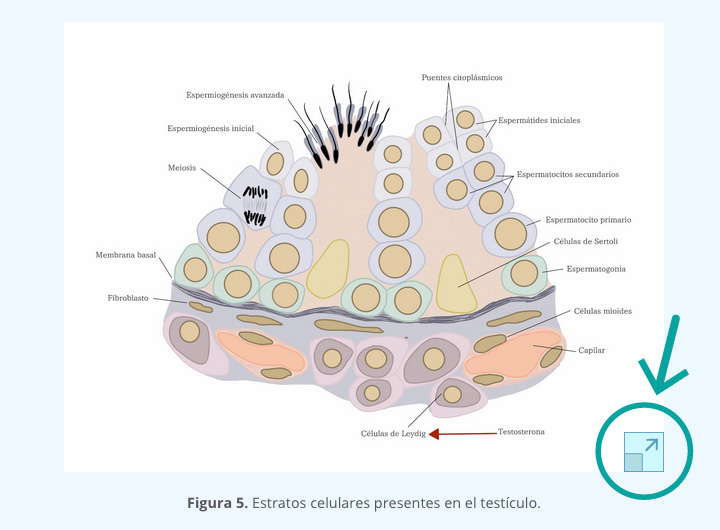
- Uno de tipo epitelial, formado por las células germinales precursoras de los gametos masculinos (espermatozoides) y femeninos (ovocitos), rodeadas de células somáticas que posteriormente se diferenciarán como células de Sertoli en el macho y células de la granulosa en la hembra.
- El otro es el tejido que forma el estroma, el cual, está constituido por tejido conectivo, vasos sanguíneos y las células precursoras de células intersticiales con actividad esteroidogénica, que son células de Leydig en el testículo y la teca interna en el ovario.
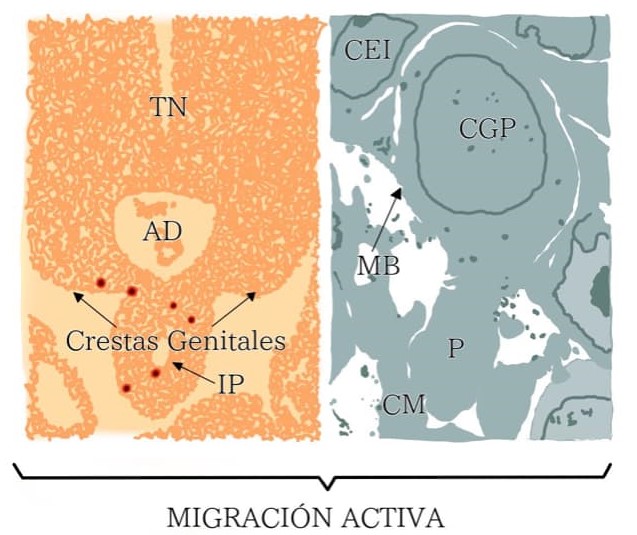
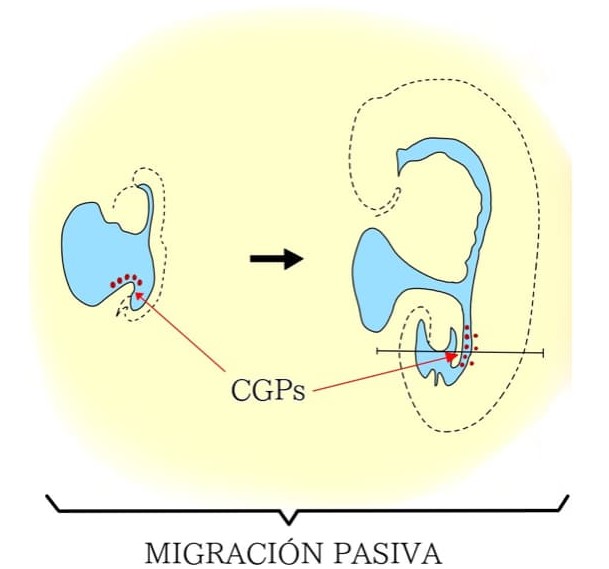
Las células germinales primordiales (CGPs) tienen un origen extragonadal y se especifican durante la gastrulación en la región posterior del epiblasto. Éste proceso es controlado por el factor de crecimiento óseo 4 (BMP4) secretado por el ectodermo extraembrionario, lo cual promueve la expresión de los factores BLIMP 1 y BLIMP14 en las células precursoras de las CGPs. Un marcador clásico para detectar a las CGPs en la base de la alantoides o en la pared del saco vitelino es la expresión de fosfatasa alcalina (FA). Las CGPs son trasladadas pasivamente desde el saco vitelino al epitelio del intestino posterior gracias a movimientos morfogenéticos, favorecidos por el continuo crecimiento del embrión. En la vecindad de la zona, donde se formarán las crestas gonadales, las CGPs abandonan el epitelio intestinal con un movimiento ameboide y se dirigen hacia las crestas a través del tejido estromático. A este tipo de migración se le conoce como migración activa (figura 2).
Aunque las células germinales primordiales son bipotenciales, expresan los genes Oct4, Nanog, Sox2, Prdm14 y Lin28, conocidos como marcadores de pluripotencia. La expresión simultánea de estos factores depende de la especie de estudio, por ejemplo, en el caso del humano los gonocitos expresan OCT4, NANOG, LIN28, pero carecen de la expresión de SOX2.
La especificación y proliferación de las CGPs son eventos que ocurren al mismo tiempo; en el caso del ratón ocurre cerca de los 7.25 dpc (días post coito). La sobrevivencia y/o proliferación de las CGPs es regulada por diversos factores de crecimiento y citocinas, que afectan positiva o negativamente a este tipo celular, entre estos factores encontramos a FGF2, FGF9, BMP4, BMP7, LIF, TGFβ y SCF.