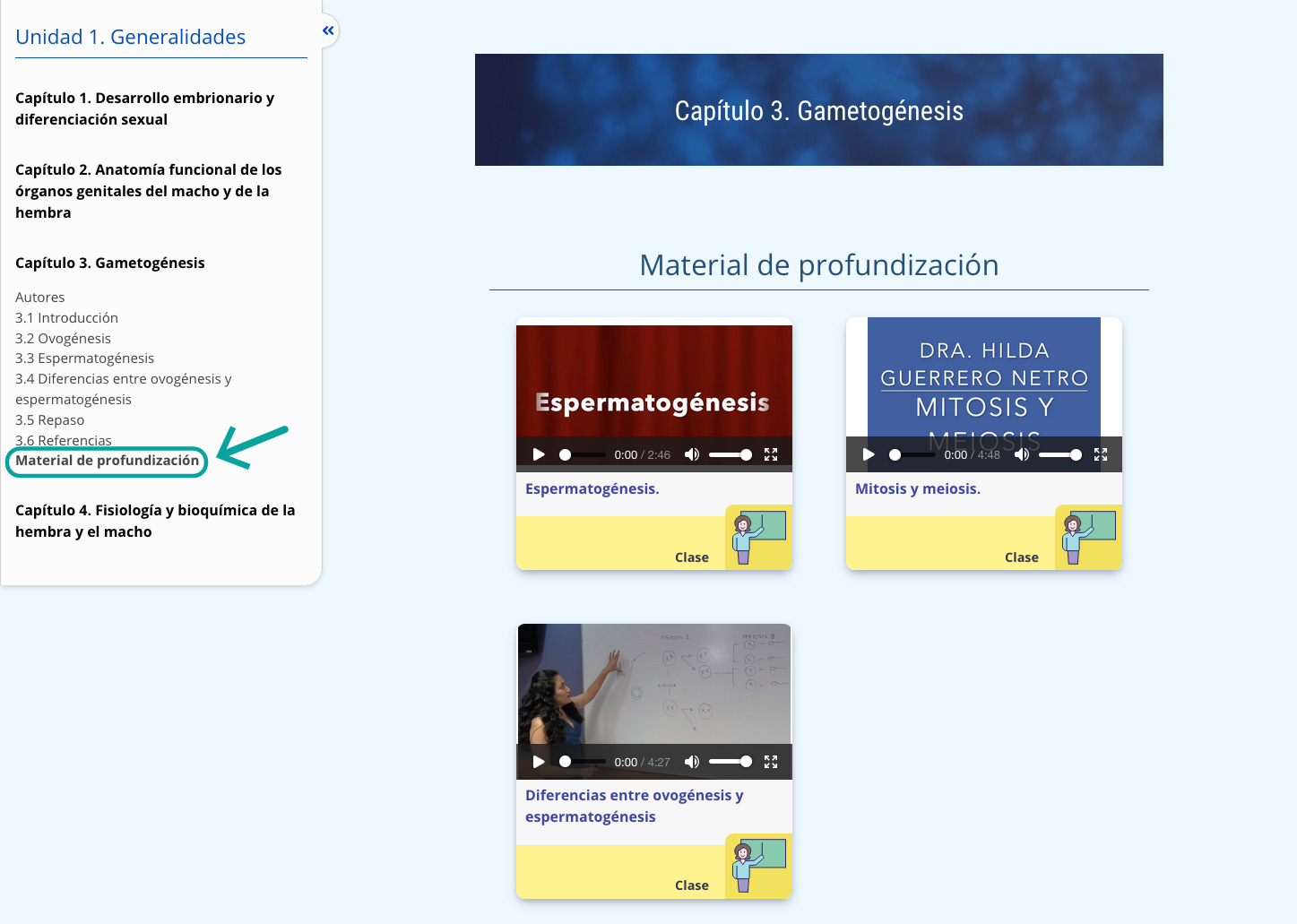
1.6 DIFERENCIACIÓN DE LOS CONDUCTOS SEXUALES
De manera simultánea al establecimiento de la gónada, se forman también a lo largo del mesonefros dos sistemas de conductos (interactivo 2).
- Los conductos mesonéfricos o de Wolff, precursores del tracto masculino: conductos deferentes, eyaculadores y vesículas seminales.
- Los conductos paramesonéfricos también conocidos como conductos de Müller, precursores del tracto femenino: trompas uterinas, útero, cérvix y la porción craneal de la vagina.
Müller
(paramesonéfricos).
(mesonéfrico).
del cuerpo uterino.
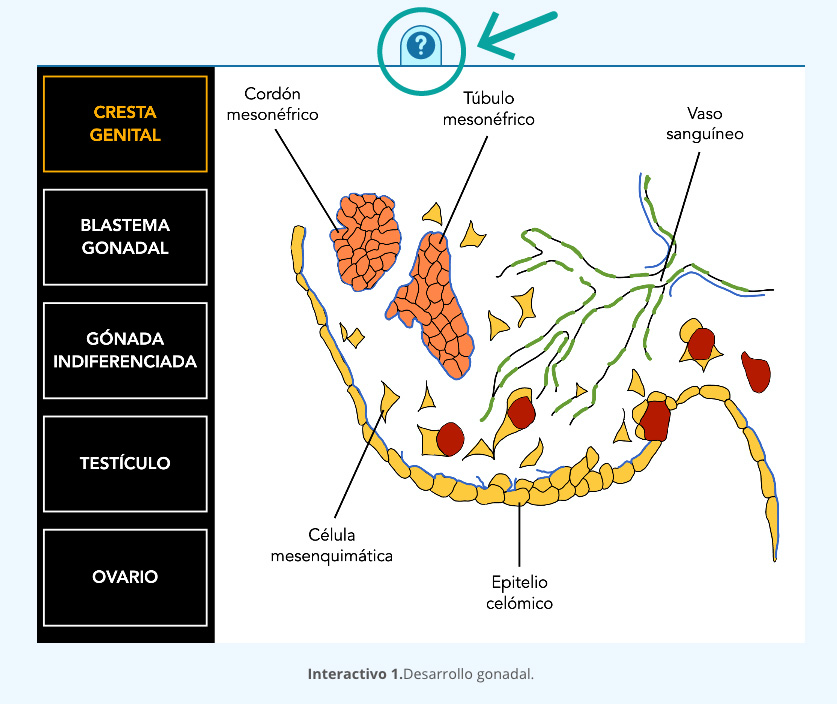
indiferenciada
(mesonefros y
gónada indiferenciada).
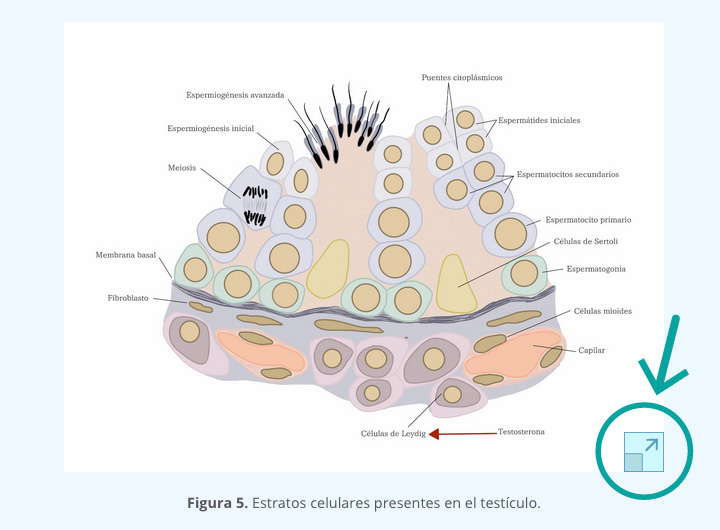
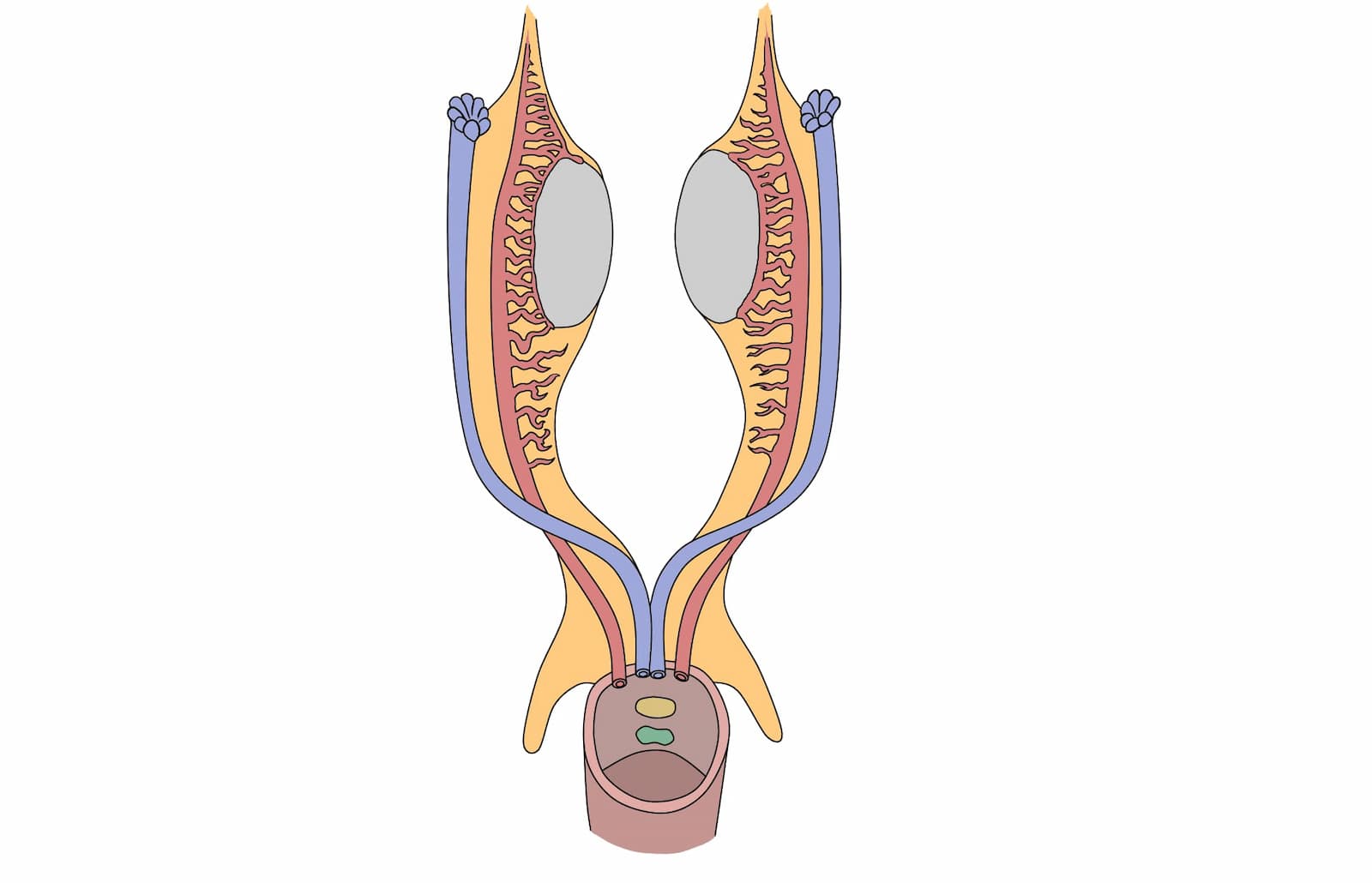
Las células del testículo fetal producen dos hormonas implicadas en la masculinización del tracto. En el estroma, las células de Leydig producen testosterona, en tanto que en los cordones testiculares las células de Sertoli secretan la AMH. Dichas hormonas contribuyen para la diferenciación de los derivados del conducto de Wolff y la regresión de los conductos de Müller, respectivamente (figura 4).
Las hormonas esteroides involucradas en la diferenciación del aparato genital masculino son los andrógenos, específicamente, la testosterona, encargada de la diferenciación del epidídimo, el conducto deferente, conducto eyaculador y las vesículas seminales. Así como la dihidrotestosterona que contribuye con la diferenciación de la próstata y los genitales externos.
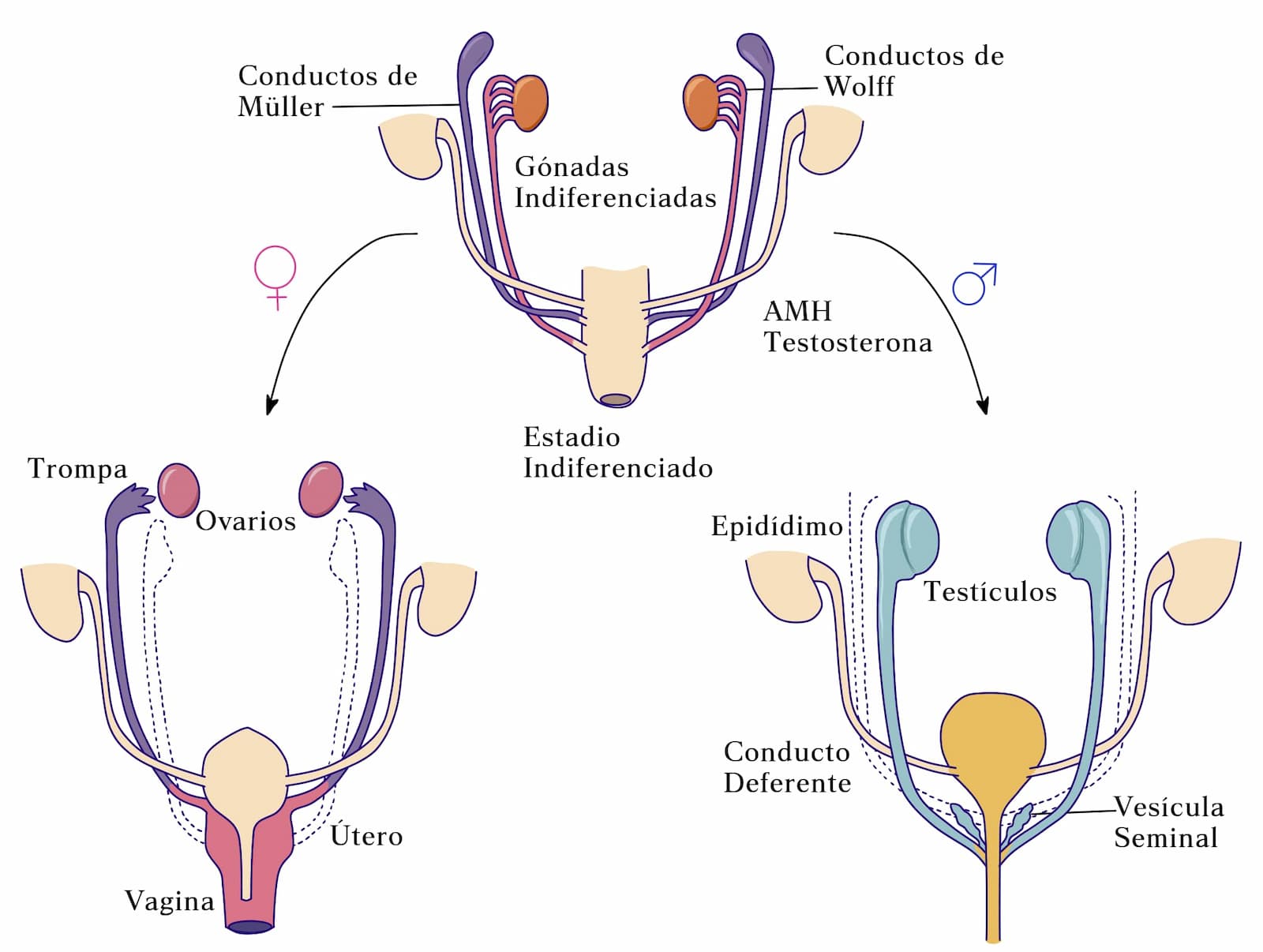
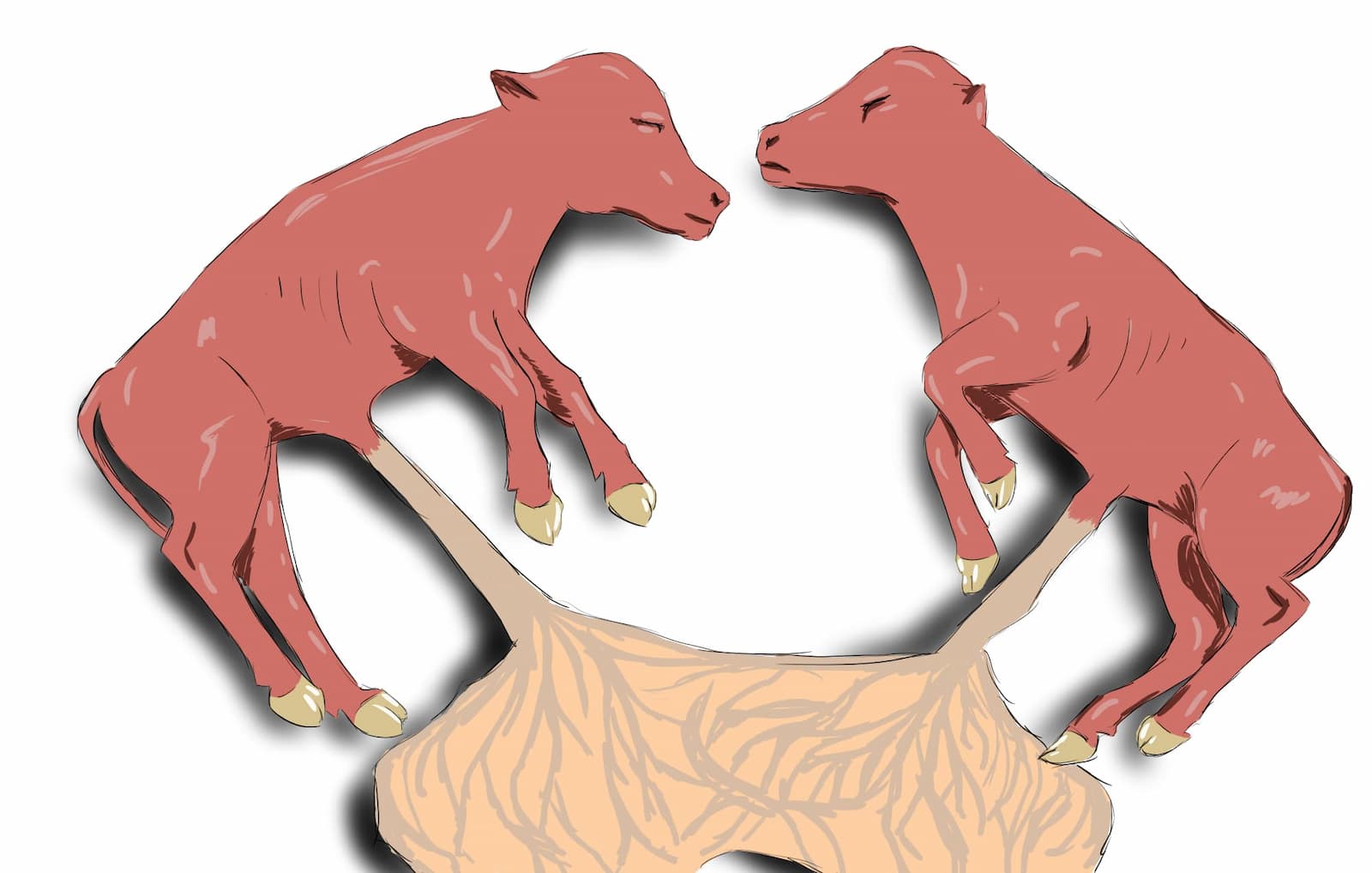
La existencia de la AMH fue propuesta por Alfred Jost, en el estudio del síndrome de Freemartin, referente a gestación gemelar de fetos con diferente sexo en bovinos. Aquí, los vasos sanguíneos de las dos placentas se anastomosan permitiendo el intercambio de sangre entre los fetos gemelos (figura 5). Al nacer, los ovarios de las hembras se encuentran atrofiados y los derivados de los conductos Müllerianos se encuentran atrofiados o inexistentes y los conductos de Wolff presentan cierto desarrollo. Este ejemplo de intersexo sirvió como modelo para postular la llamada “teoría de la base hormonal de la diferenciación sexual fenotípica”, basada en la producción dual de hormonas que controlan la regresión de los conductos de Müller y la diferenciación del tracto masculino. Alfred Jost sugirió la existencia de un factor inhibidor de los conductos Müllerianos producido por los testículos del co-gemelo.
La AMH es una glicoproteína perteneciente a la familia TGFβ y es uno de los primeros marcadores de diferenciación de las células de Sertoli. La secreción de AMH continúa durante la vida fetal y posnatal, declinando en la pubertad. La permanencia de la expresión de AMH en el testículo adulto se considera un signo de inmadurez de las células de Sertoli. La expresión de la AMH en el testículo fetal es regulada directamente por Sox9, Sf1, Wt1 y Dax1. En el caso del ovario la expresión de AMH es indetectable en el ovario embrionario. Sin embargo, su expresión es evidente a partir de la formación de los folículos primarios y es considerado un marcado de la reserva folicular.
Durante la vida embrionaria y fetal los ovarios no expresan AMH y esto permite la diferenciación de los conductos de Müller. Además, el patrón de diferenciación requiere de la expresión de una compleja cascada de expresión génica donde los genes Hox tienen un papel central. Hoxa9 es expresado para especificar los oviductos, Hoxa10 se expresa en el útero, Hoxa11 en la parte posterior del útero y el cérvix y Hoxa13 en la porción superior de la vagina. En tanto que la expresión de los genes Wnt tiene una función importante en mantener la expresión de los genes Hox mencionados.