4.2 FISIOLOGÍA DE LA HEMBRA
4.2.1 INTRODUCCIÓN
La reproducción en la hembra es un evento complejo que involucra no solo al aparato reproductor (hormonas ováricas), si no también al cerebro (hipotálamo y gonadotropinas hipofisarias) y su capacidad para responder a diferentes niveles hormonales, estímulos externos como las horas luz y a la condición corporal del animal. Todos estos factores regulan la producción de estrógenos, los cuales determinan cuándo se da el inicio de la pubertad, la actividad cíclica del ovario, la gestación y el parto.
4.2.2 CONTROL NEUROENDOCRINO DEL CICLO ESTRAL
La reproducción de los animales domésticos es un proceso que involucra al eje hipotalámico hipofisario gonadal, lo cual da como resultado una compleja señalización dentro del cerebro que posteriormente se transmite al ovario y resulta en la ovulación. Esta señalización comienza con la producción de la hormona liberadora de gonadotropinas (GnRH), una hormona polipeptídica (constituida por 10 aminoácidos) producida por neuronas del hipotálamo, actualmente se considera que hay dos o más tipos de GnRH, incluyendo la GnRH I la cual tiene función dentro del hipotálamo y cuya estructura varía entre especies; la GnRH II, tiene función extra hipotalámica actuando como regulador y con forma altamente conservada entre especies.
La GnRH tienen una vida media de 10 minutos y se libera en forma de pulsos en el torrente sanguíneo portal hipofisario, la sangre lleva a la GnRH a la glándula hipófisis donde su forma de secreción pulsátil determina la liberación en fases de la hormona folículo estimulante (FSH) y la hormona luteinizante (LH), lo cual resulta en la presentación de los ciclos estrales característicos de las hembras (figura 1).

Estas dos hormonas están clasificadas en el grupo de glicoproteínas, esto se refiere a proteínas (cadenas largas de aa) unidas covalentemente a carbohidratos en algunos de sus aa. Dentro de esta misma familia encontramos a la gonadotropina coriónica humana (hCG) y la gonadotropina coriónica equina (eCG). Todas éstas están formadas por dos subunidades expresadas en genes distintos, la alfa (idéntica para todas las glicoproteínas de una especie animal) y la beta (específica para cada hormona). Hablando meramente de la hCG, tiene función de FSH en el humano, sin embargo, si se usa en otras especies tendrá funciones que cubren las de LH; mientras que la eCG producida en las copas endometriales, actúa como LH dentro del équido, mientras que al usarse en otras especies funge como FSH.
4.2.3 FASES DEL DESARROLLO FOLICULAR Y ESTEROIDOGÉNESIS
En los animales domésticos, los ciclos estrales de las hembras están caracterizados por la presencia de oleadas foliculares, cada ciclo estral puede presentar de dos a tres oleadas foliculares (figura 2).

Los folículos son pequeños sacos presentes en el ovario, los cuales contienen las células de la teca, granulosa y el ovocito (figura 3) (consultar capítulo 3 "Gametogénesis").
.jpg)
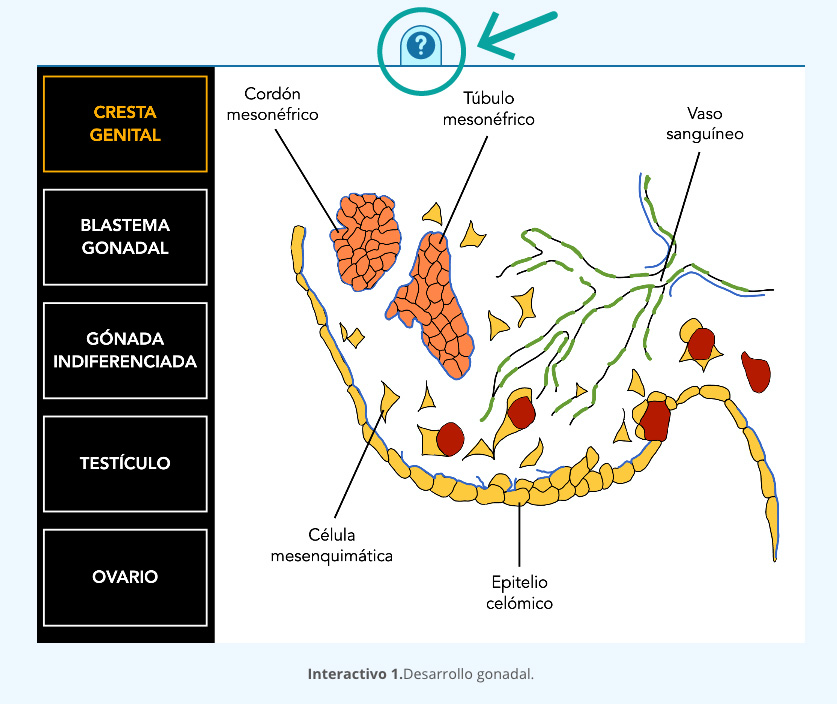
Para que la ovulación se lleve a cabo, el folículo debe desarrollarse junto con las células en su interior, este evento folicular consta de tres etapas: reclutamiento, selección y dominancia (interactivo 1). El reclutamiento inicia con la llegada de la FSH al ovario, que como su nombre lo indica, estimula el crecimiento de folículos que se encuentran en la etapa tónica, es decir, dependientes de gonadotropinas. Durante el reclutamiento varios folículos recibirán el estímulo de la FSH y comenzarán su crecimiento (figura 4).
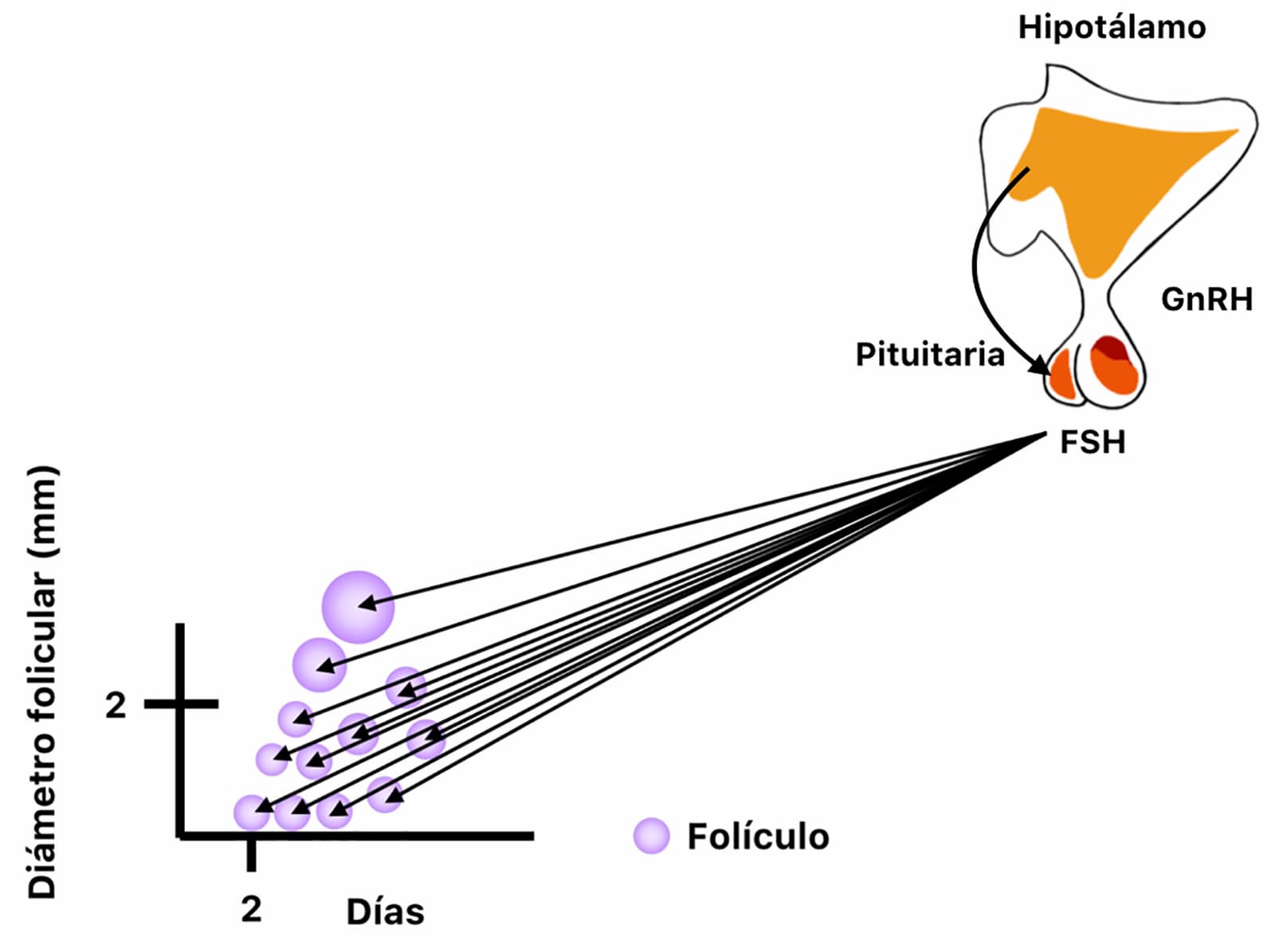
Adicionalmente, las células dentro del folículo comenzarán el proceso de esteroidogénesis, en lo que se conoce como “La teoría de las dos células para la producción de estrógenos”.
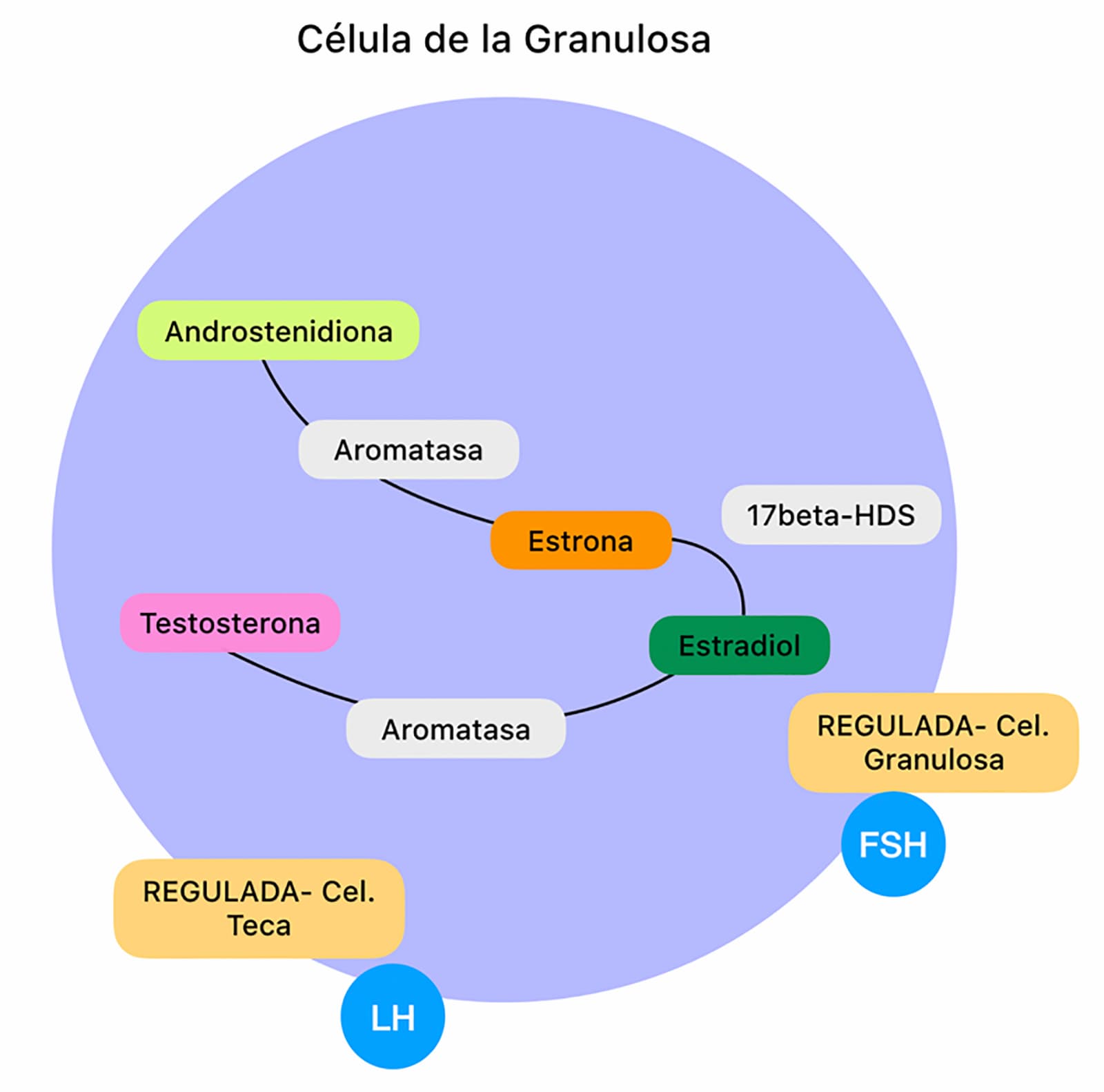
En esta teoría se establece que para la producción de estradiol se necesitan dos tipos celulares, las células de la teca y las células de la granulosa. La esteroidogénesis (proceso por el cual se sintetizan hormonas esteroideas a partir del colesterol) inicia con el colesterol en las células de la teca y termina con diferentes esteroides incluyendo el estradiol en las células de la granulosa. La producción de esteroides está mediada por enzimas que son responsables de transformar un esteroide a otro.
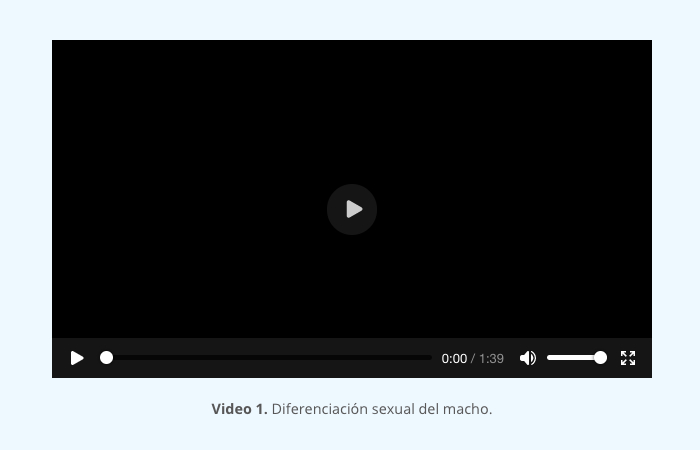
El colesterol entra a las células de la teca por medio de lipoproteínas de baja densidad; una vez dentro de la célula éste se mantiene en gotas lipídicas en forma de ésteres de colesterol los cuales se transforman por medio de la hidrolasa del éster del colesterol en colesterol libre; éste es altamente hidrofóbico lo cual ayuda a metabolizarse e interiorizarse en la mitocondria. Esta internalización está mediada por la enzima StAR (Steroidogenic acute regulatory protein) y una vez dentro de la mitocondria, el colesterol es convertido en pregnenolona (primer esteroide) por medio de P450scc (cytocrhrome P450 cholesterol side-chain cleavage). La pregnenolona, a su vez, es convertida por medio de la 3β-hydroxysteroid dehydrogenase (3β-HSD) a progesterona, ésta puede pasar a las células de la granulosa o seguir en las de la teca donde se convierte a androstenediona por medio de la P45017-OH (enzyme cytochrome P450 17α-hydroxylase) y 3β-HSD. Las células de la teca convierten una pequeña cantidad de androstenediona a testosterona por medio de la 17β-HSD (17β-hydroxysteroid dehydrogenase) (video 1).
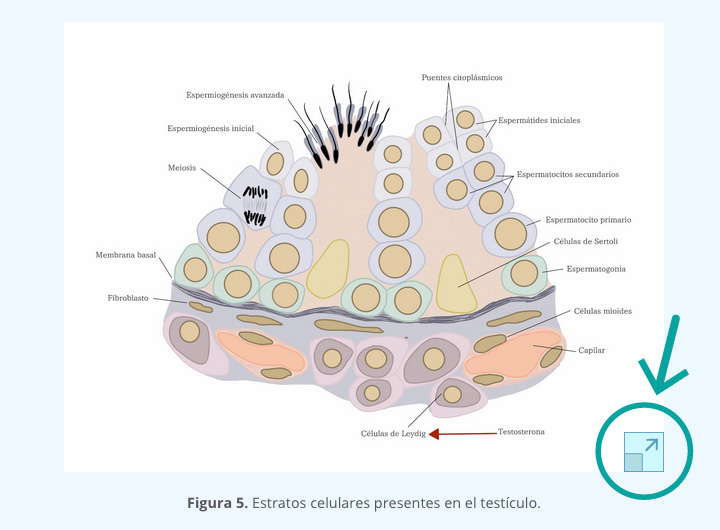
Tanto testosterona y androstenediona se convierten a estradiol en las células de la granulosa; la androstenediona se transforma en estrona por medio de la aromatasa y de estrona a estradiol por medio de 17β-HSD, mientras que la testosterona se convierte directamente a estradiol por medio de la aromatasa. Esta producción de estradiol en folículos pequeños está regulada por la FSH en las células de la granulosa y por la LH en las células de la teca (figura 5).

Para la última etapa del desarrollo folicular, la dominancia, se debe producir un pico de LH el cual induce la ovulación y para el cual las células de la granulosa deben adquirir los receptores. Una vez que el folículo alcanza la etapa de pre-ovulatorio, las células de la granulosa adquieren los receptores para LH, sin embargo, el pico de LH está regulado por la presencia del cuerpo lúteo, es decir, mientras esté presente la estructura produciendo progesterona, el pico de LH no tiene lugar y por lo tanto aunque un folículo sea seleccionado y esté listo para la ovulación, si no hay pico de LH, éste se vuelve atrésico e inicia una nueva oleada folicular (figura 7). Sin embargo, una vez que se lleva a cabo la regresión el cuerpo lúteo, la falta de progesterona permite el pico de LH y con esto se induce la ovulación del folículo dominante de la oleada folicular en curso (figura 8).


4.2.4 REGULACIÓN DE LA GnRH POR MEDIO DE LA KISSPEPTINA
La hipótesis de la masa grasa crítica (critical fat mass hypothesis) establece que la reproducción es una función que demanda energía y por lo tanto se encuentra limitada a su disponibilidad, además de estar influenciada por ambos extremos de la nutrición (la obesidad y la desnutrición), alterando, por ejemplo, la edad en la que se presenta la pubertad. Este control de la reproducción por medio del estado nutricional es el resultado de la acción de diferentes hormonas que detectan el estado metabólico del animal y transmiten esta información al eje hipotalámico encargado de la producción de GnRH.
Entre las señales involucradas se encuentra la leptina, hormona proveniente de los adipocitos, la cual está presente en el organismo de manera proporcional al tamaño de los reservorios de grasa, considerada esencial para la reproducción. La leptina es considerada un indicador de suficiencia energética, permitiendo a diferentes sistemas detectar las reservas de energía. Una baja cantidad de leptina resulta en anormalidades en el funcionamiento de diferentes sistemas, incluyendo el aparato reproductor.
A pesar de esto, su sitio de acción a nivel hipotalámico no es claro, una teoría indica que la leptina regula directamente la producción de GnRH, se ha observado en casos de obesidad que altos niveles de leptina reducen las funciones gonadales, sin embargo, las neuronas encargadas de la producción de GnRH parecen no tener receptores de leptina o, en especies donde se encuentran presentes como el ratón, la inhibición de estos receptores parece no afectar la reproducción. Actualmente se cree que la leptina tiene acción sobre el gen Kiss-1 el cual codifica para la kisspeptina.
La kisspeptina es una hormona proteica de 145 aa, es secretada por neuronas localizadas en el hipotálamo, su posición con referencia a las neuronas que producen la GnRH varía por especie, sin embargo, en roedores las neuronas encargadas de producir kisspeptina se encuentran en el área pre-óptica y el núcleo del infundíbulo (POA) mientras que las de GnRH se encuentran en la eminencia media (ME) (figura 9).

A diferencia de la leptina, las neuronas encargadas de la secreción de GnRH poseen receptores específicos para la kisspeptina por medio de los cuales se regula la liberación de GnRH a la circulación y con esto, el ciclo estral de un animal. A pesar de que la GnRH podría considerarse la principal reguladora del ciclo estral, el descubrimiento de la kisspeptina como regulador clave de la GnRH ha llevado a una nueva etapa en el entendimiento de la regulación neuroendocrina de la reproducción.
4.2.5 ROL DE LA MELATONINA EN LA REPRODUCCIÓN
En especies estacionales como los equinos y pequeños rumiantes, la actividad ovárica está regulada por el fotoperiodo, es decir la cantidad de horas luz a la que están expuestas. Estos animales responden a los cambios en la duración del día y regulan la función ovárica a través de la melatonina, la cual es producida por la glándula pineal durante la noche, está considerada dentro del grupo de las indolaminas, moléculas derivadas de un aa modificado, en este caso el triptófano; estas moléculas, al ser hidrosolubles, tienen receptores en la membrana citoplasmática de la célula blanco. Regula la secreción de GnRH provocando una sub-regulación o sobre-regulación (dependiendo de la especie) de la FSH y LH. En equinos, especie estacional de días largos, la melatonina sub-regula la secreción de la GnRH, es decir, durante el invierno, la exposición a menos horas luz resulta en un aumento de melatonina, la cual disminuye la secreción de GnRH lo que conlleva a una menor secreción de FSH y LH (figura 10). Mientras que los pequeños rumiantes, especies con etapa reproductiva durante el invierno, el aumento de melatonina resulta en una mayor producción de GnRH y por lo tanto de FSH y LH (figura 11).


En especies no estacionales como los bovinos, el descubrimiento de receptores de melatonina MT1 y MT2 en células de la granulosa, cumulus y ovocitos parece indicar un posible rol en la reproducción a través de un efecto directo sobre la maduración del ovocito, sin embargo, no hay estudios suficientes para comprobar esta hipótesis.