7.2 TRANSPORTE DE LOS GAMETOS
7.2.1 TRANSPORTE DEL ÓVULO
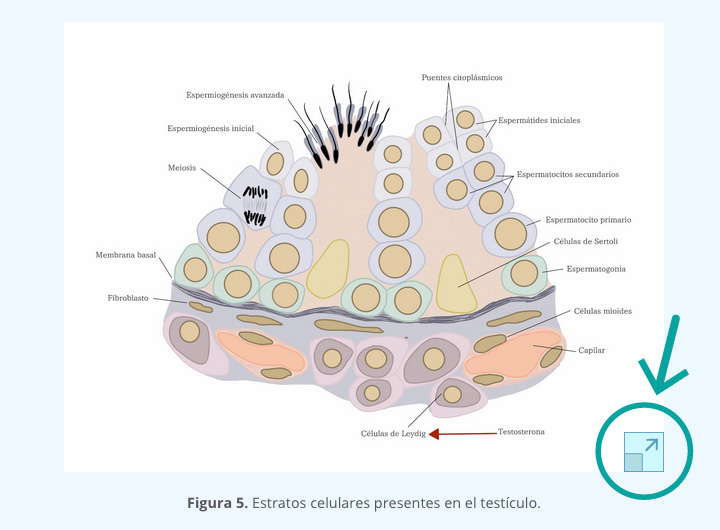
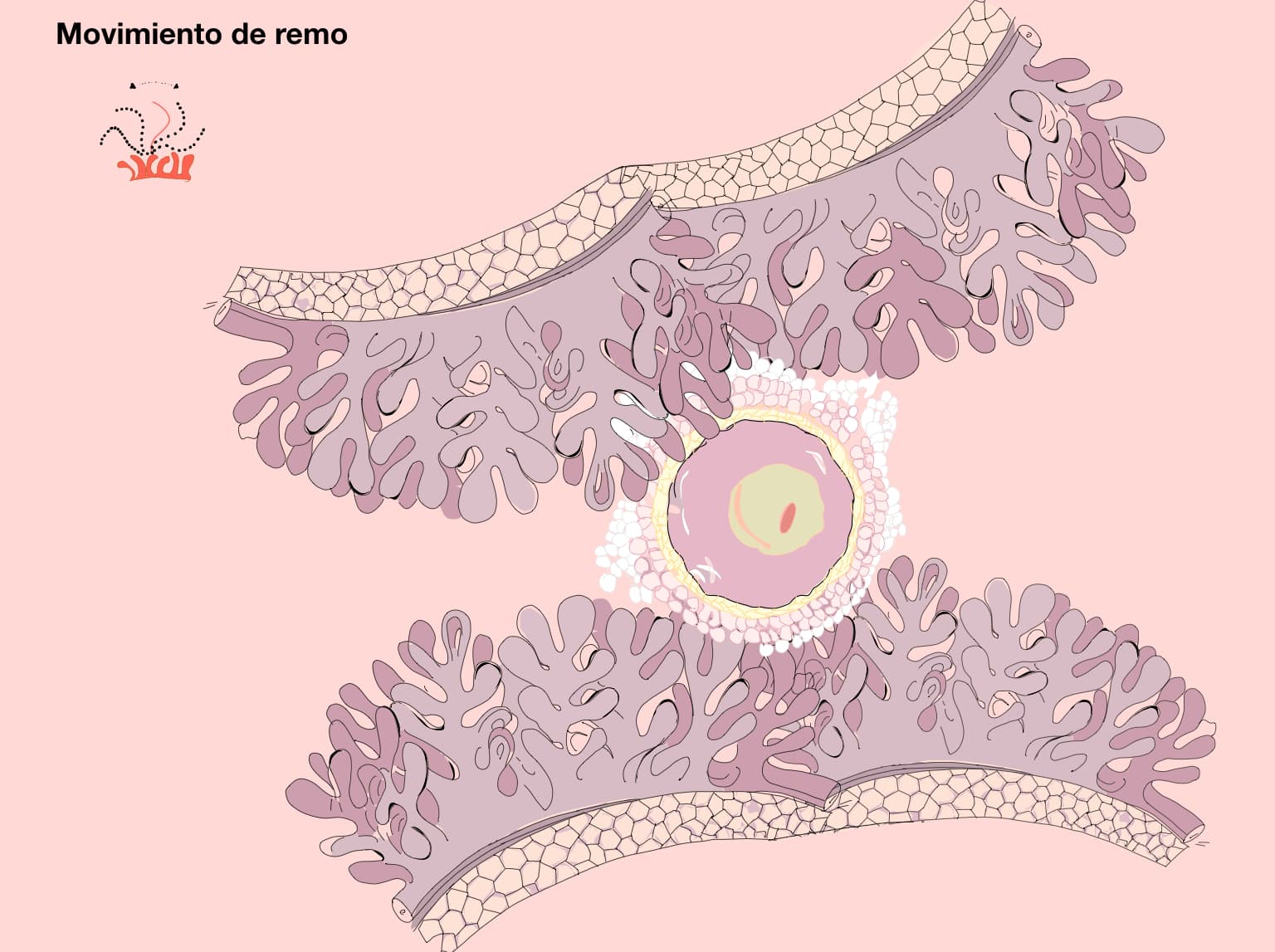
Al salir del ovario, el óvulo es una célula altamente diferenciada y se encuentra rodeada por un conjunto de células foliculares que forman dos arreglos: el cúmulo ovígero y la corona radiada (en contacto con la zona pelúcida); estas estructuras en conjunto con el óvulo reciben el nombre de complejo ovocito-células del cumulus (COC). El infundíbulo, mediante la acción vigorosa de los cilios de la fimbria, atrapa al óvulo para conducirlo hacia su lumen; su captura es facilitada por la presencia del cúmulo ovígero ya que le proporciona una mayor superficie sobre la cual los cilios pueden actuar. El COC es de suma importancia para el transporte normal del óvulo por la región ampular del oviducto que se da por el movimiento de los cilios epiteliales y por la contracción de las células musculares del mismo (figura 1). Ambos movimientos dependen de la presencia de estrógenos y de progesterona.
Según sea la especie, el transporte por la ámpula puede tomar de unos pocos minutos a varias horas; aunque, en la mayoría de las especies el transporte por todo el oviducto toma aproximadamente tres días, esté o no fertilizado el óvulo. El caso de la yegua constituye una excepción, puesto que en ella el óvulo no fecundado puede permanecer por semanas o meses en el oviducto.
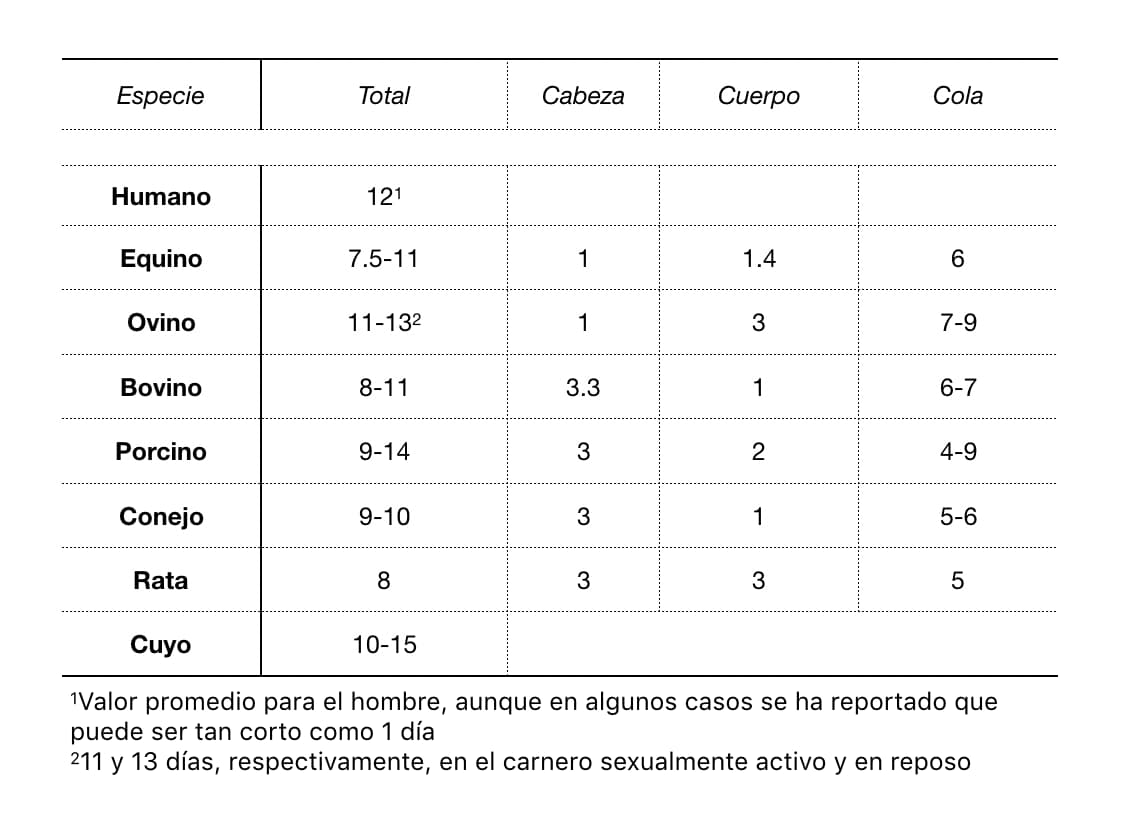
Debido a que en la mayoría de las especies la receptividad sexual se inicia varias horas antes de la ovulación, cuando el óvulo llega al sitio de fertilización, los espermatozoides ya se encuentran ahí. La vida fértil del óvulo es relativamente corta; en caso de no ser fertilizado puede morir después de 6 a 24 horas (cuadro 1). Después de unas horas empieza su envejecimiento, cambio que puede afectar seriamente el desarrollo embriológico posterior.
El transporte del óvulo puede fallar en diferentes puntos:
- Puede quedarse en la superficie del ovario, y si es fertilizado allí, se desarrolla una gestación ectópica que es sumamente rara en los animales domésticos.
- Puede caer en la cavidad peritoneal, generalmente después de su fertilización en el ovario e intenta desarrollarse adosado a uno de los órganos abdominales (gestación abdominal).
- El óvulo fecundado se queda en el oviducto en lugar de migrar hacia el útero (gestación tubárica).

7.2.2 TRANSPORTE DE LOS ESPERMATOZOIDES
Los espermatozoides liberados del testículo son morfológicamente maduros, pero son funcionalmente incompetentes debido a que carecen de capacidad de movimiento progresivo y de la habilidad para unirse a la zona pelúcida; para que los espermatozoides de mamíferos presenten esta habilidad, se deben llevar a cabo dos fases distintas llamadas maduración (en el epidídimo) y capacitación (en aparato reproductor de la hembra) (interactivo 1).
Los espermatozoides deben pasar por las tres porciones del epidídimo (cuadro 2). En todas las especies, los espermatozoides tomados directamente de la porción anterior de la cabeza del epidídimo son incapaces de fecundar a un óvulo recién liberado, pero su capacidad aumenta en porcentaje conforme se avanza a lo largo del órgano hasta su porción posterior (rata: 27%; hamster 47%; conejo 68-71%; carnero 33%; cerdo 46%) y es siempre máxima en los espermatozoides tomados de la cola del epidídimo.
La adquisición de la motilidad progresiva es una de las modificaciones que ocurren a los espermatozoides durante su paso por cada una de las regiones del epidídimo. Estructuralmente se han observado dos rasgos de la maduración espermática: la remodelación del acrosoma y la migración de la gota citoplasmática.
Los espermatozoides pueden ser emitidos al exterior durante la cópula, la masturbación o en emisiones espontáneas. En ausencia de estos eventos, hay un flujo continuo de espermatozoides por la uretra, puesto que son arrastrados por la orina al exterior. Durante el proceso de la emisión seminal y de la eyaculación, los espermatozoides maduros, suspendidos en las secreciones del testículo y del epidídimo, se mezclan antes de llegar a la uretra con las secreciones de las glándulas accesorias; en conjunto constituye el semen.
El líquido seminal sirve de vehículo pero también proporciona sustancias necesarias para el metabolismo energético del espermatozoide. Cabe señalar que si un animal tiene una intensa actividad sexual, es decir, eyacula muy frecuentemente, aumentará la proporción de espermatozoides con gota citoplasmática en los eyaculados subsecuentes, lo que es indicativo de que no se completa el proceso de maduración.
Durante la cópula, el semen puede ser depositado en la vagina (vaca, oveja, coneja, primates, gata) o directamente dentro del útero (yegua, cerda, camélidos y roedores). En el caso de los roedores la formación de un tapón vaginal gelatinoso, inmediatamente después del coito, impide la salida del semen de la vagina. Así mismo, en el cerdo la porción gelatinosa del eyaculado funciona de manera similar. El sitio de depósito del semen en el tracto de la hembra definirá la distancia y las barreras anatómicas que el espermatozoide encontrará durante su transporte hacia el oviducto. Estas barreras, además de restringir el paso de los espermatozoides, establecerán un gradiente espermático en el tracto de la hembra y funcionarán como un reservorio de espermatozoides. El establecimiento del gradiente espermático posiblemente reduce el riesgo de poliespermia (figura 2).
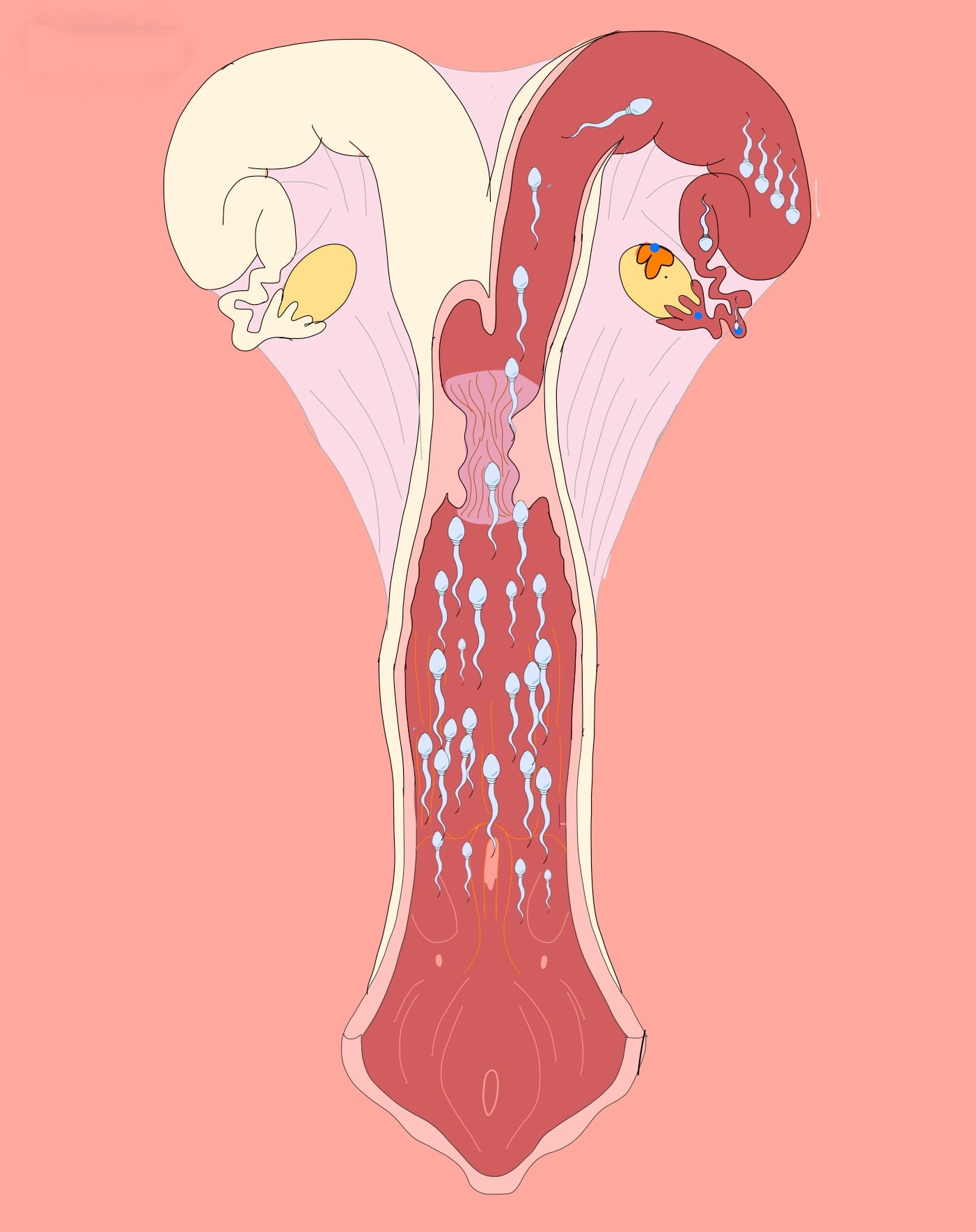
Cabe señalar que la distribución de los espermatozoides en el tracto de la hembra se da por dos tipos de transporte:
- Fase rápida: evento pericoital caracterizado por la presencia de espermatozoides en el oviducto a los pocos minutos después de la monta; atribuido a la contracción muscular del tracto reproductor de la hembra. Sin embargo, estos espermatozoides no forman parte de la población fertilizante de espermatozoides que se establece en el oviducto, ya que la mayoría de estos se encuentran débiles, muertos o dañados.
- Fase prolongada de migración: se establecen los gradientes espermáticos, los reservorios espermáticos y por consiguiente, contiene a los espermatozoides competentes que pueden llegar al oviducto.
Los espermatozoides competentes que atraviesan la unión útero tubárica, previo a la ovulación, se acumulan en la parte baja del istmo y permanecen ahí hasta el inicio de la ovulación (figura 3). La parte baja del istmo no sólo funciona como un reservorio espermático, también regula el ascenso espermático hacia la ámpula. De hecho, la interacción funcional entre los espermatozoides competentes y los fluidos luminales, junto con la superficie epitelial del tracto reproductor de la hembra, promueven la selección de las poblaciones espermáticas fisiológicamente normales.
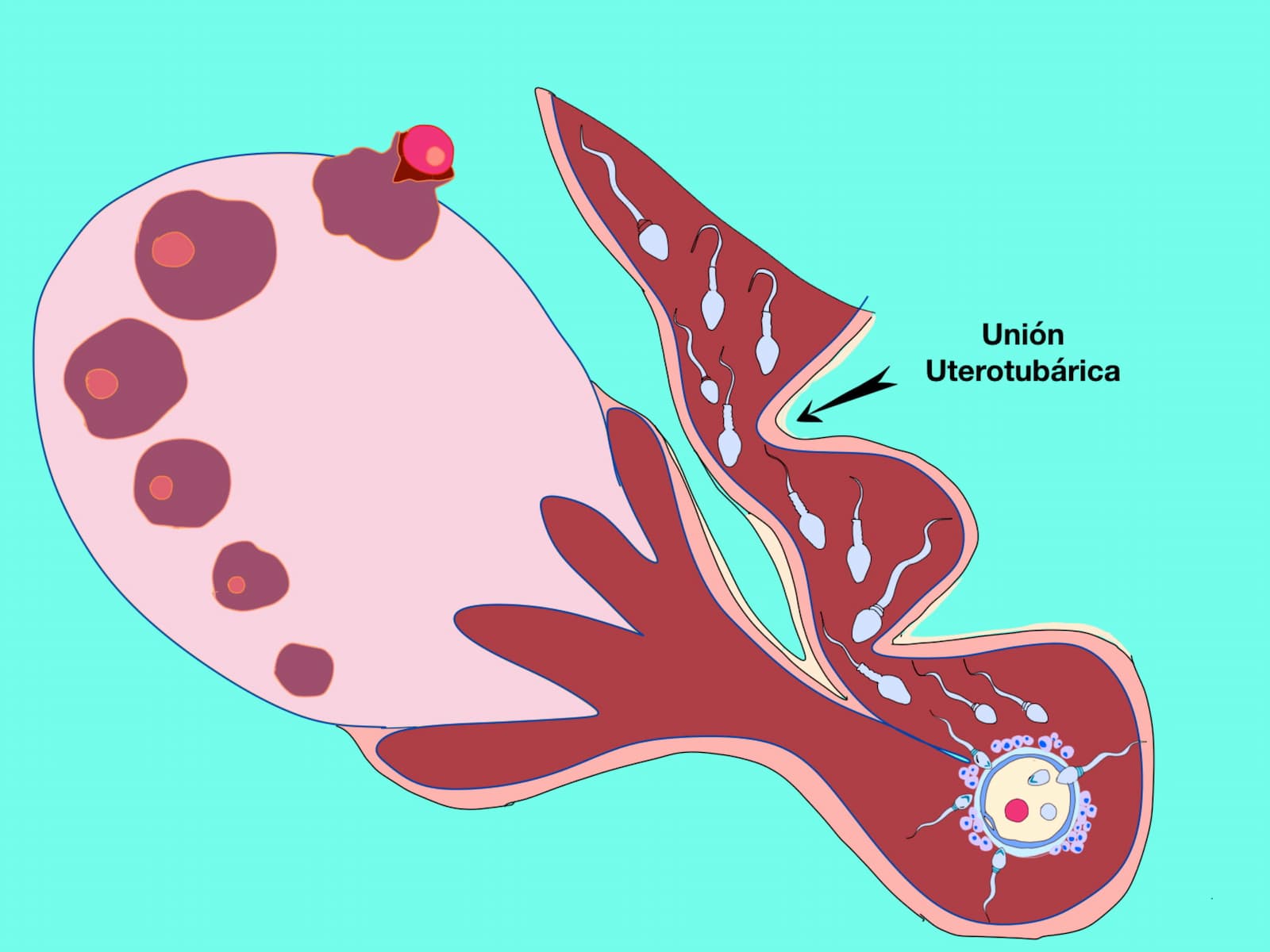
Por otra parte, diversas líneas de evidencias involucran a la proteína Adam 3 (por sus siglas en inglés: a disintegrin and metallopeptidase domain; un dominio de desintegrina y metalopéptidasa) como un importante factor espermático para la migración de los espermatozoides al oviducto. En el caso del bovino, se ha sugerido que en la superficie espermática la proteína de aglutinación (BSP: binder of sperm protein) y proteínas de la familia de las anexinas en la superficie de las células epiteliales del oviducto juegan un papel importante en el almacenamiento de los espermatozoides en la región del istmo.
La motilidad espermática puede ser requerida para que los espermatozoides migren a través de la unión útero tubárica, pero es indispensable para que asciendan a la ampolla y penetren en las envolturas del óvulo. En el tracto de la hembra, la motilidad espermática es modulada por medidas impuestas en la dinámica del flagelo, ya sea espacialmente por factores limitantes o por las características de la superficie epitelial o de los fluidos secretados.
Estudios teóricos recientes por computadora han sugerido que el incremento de la viscosidad de un fluido puede causar que el flagelo espermático modifique su patrón de movimiento, por lo que a pesar de que este tipo de fluidos opongan resistencia al movimiento progresivo, el espermatozoide de los mamíferos debe ser capaz de migrar en fluidos viscosos fisiológicos con una velocidad progresiva similar a la que realiza en los fluidos salinos; en gran medida, dicho incremento en la viscosidad debe resultar en un aumento de los requerimientos metabólicos, los cuales deben ser proporcionados por la glucólisis (figura 4).
.jpg)
.jpg)
Aún cuando la vida fértil del espermatozoide es muy breve en la mayoría de las especies (cuadro 1), hay algunas excepciones. Por ejemplo, en las gallinas, los espermatozoides son almacenados en unas estructuras crípticas dentro del oviducto y son liberados gradualmente conforme pasan los óvulos para ser fertilizados. Los espermatozoides pueden conservarse hasta por 3 semanas o más. En algunos murciélagos el coito ocurre en el otoño, posteriormente los animales hibernan durante 3-4 meses y ovulan al despertar cuando ya es primavera, de esta manera los óvulos son fertilizados por los espermatozoides que fueron depositados en el otoño anterior.
De los millones de espermatozoides eyaculados o inseminados sólo unos pocos llegan a alcanzar el oviducto (cuadro 3); la relación de espermatozoides-ovocitos en el sitio de fertilización debe ser 1:1. Los espermatozoides deben recorrer una distancia considerable desde su liberación del testículo hasta el sitio de fertilización, por lo que existe un gran número de espermatozoides que se desperdicia. De igual manera, muchos de los espermatozoides son rápidamente eliminados en la parte baja del aparato reproductor de la hembra por un flujo retrógrado del cérvix a la vagina (figura 5).

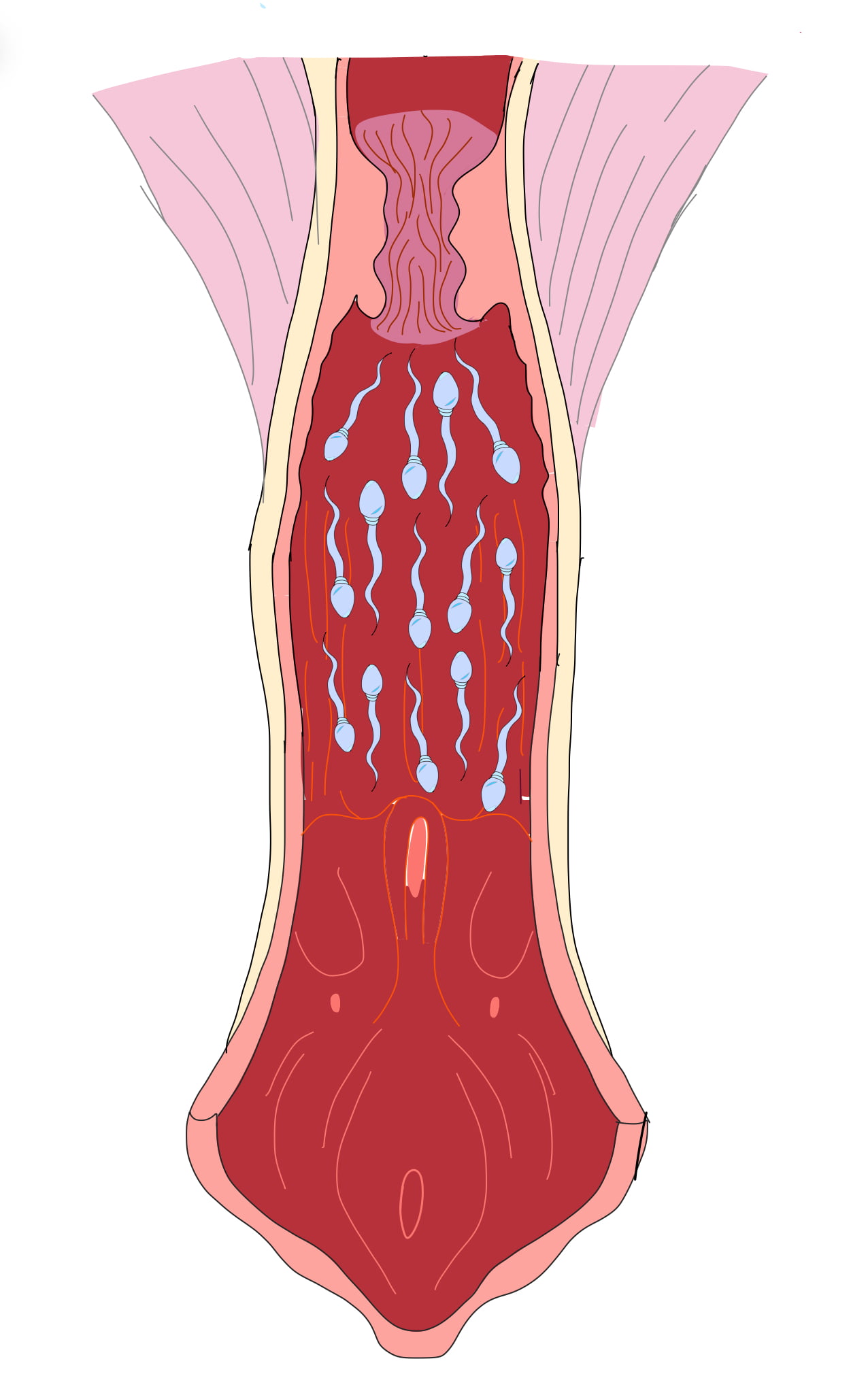
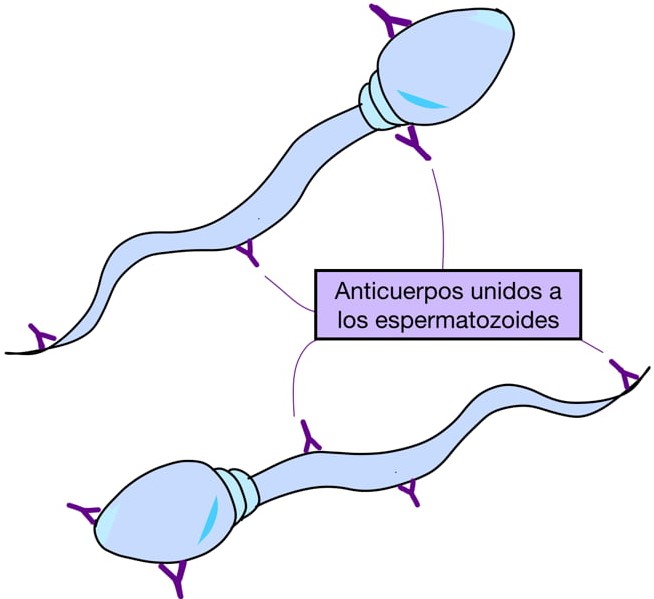
Adicionalmente, una inflamación fisiológica ocurre en respuesta a la inseminación, por lo que también se lleva a cabo una fagocitosis de los espermatozoides. A veces, algunos espermatozoides duran más tiempo en el útero y cuando llega el embrión son fagocitados por las células del trofoblasto. También se puede presentar una reacción antigénica en la hembra e inclusive es posible detectar anticuerpos anti-espermatozoides tanto en la sangre como en las secreciones del conducto genital de hembras servidas (figura 6).