7.4 FERTILIZACIÓN
La interacción del espermatozoide y del óvulo inicia una serie de transformaciones que involucran a los componentes nucleares y citoplásmicos de ambos gametos. Estas transformaciones constituyen el proceso de fertilización que comienza con la interacción y subsecuente fusión de los gametos y termina con la asociación de los grupos correspondientes de cromosomas derivados de los dos pronúcleos, uno de origen materno y el otro paterno. Con la fertilización se decide el sexo genético del individuo, dependiendo del tipo de espermatozoide (X o Y) que haya fertilizado el óvulo. Los aspectos esenciales de la fertilización son: la asociación del genoma paterno con el materno y la activación del óvulo.
Durante el crecimiento del ovocito la membrana plasmática adquiere la capacidad para la fusión. Se sabe que los ovocitos pequeños de los folículos primordiales no son capaces de fusionarse con el espermatozoide, pero al alcanzar un diámetro de aproximadamente 20 μm, mientras el ovocito está aún latente en la profase de la primera división meiótica, la fusión puede ocurrir. Durante la fertilización normal in vivo el ovocito no conocerá al espermatozoide hasta la ovulación en la que tendrá un diámetro de 80 μm y se quedará en la metafase II.
7.4.1 PENETRACIÓN
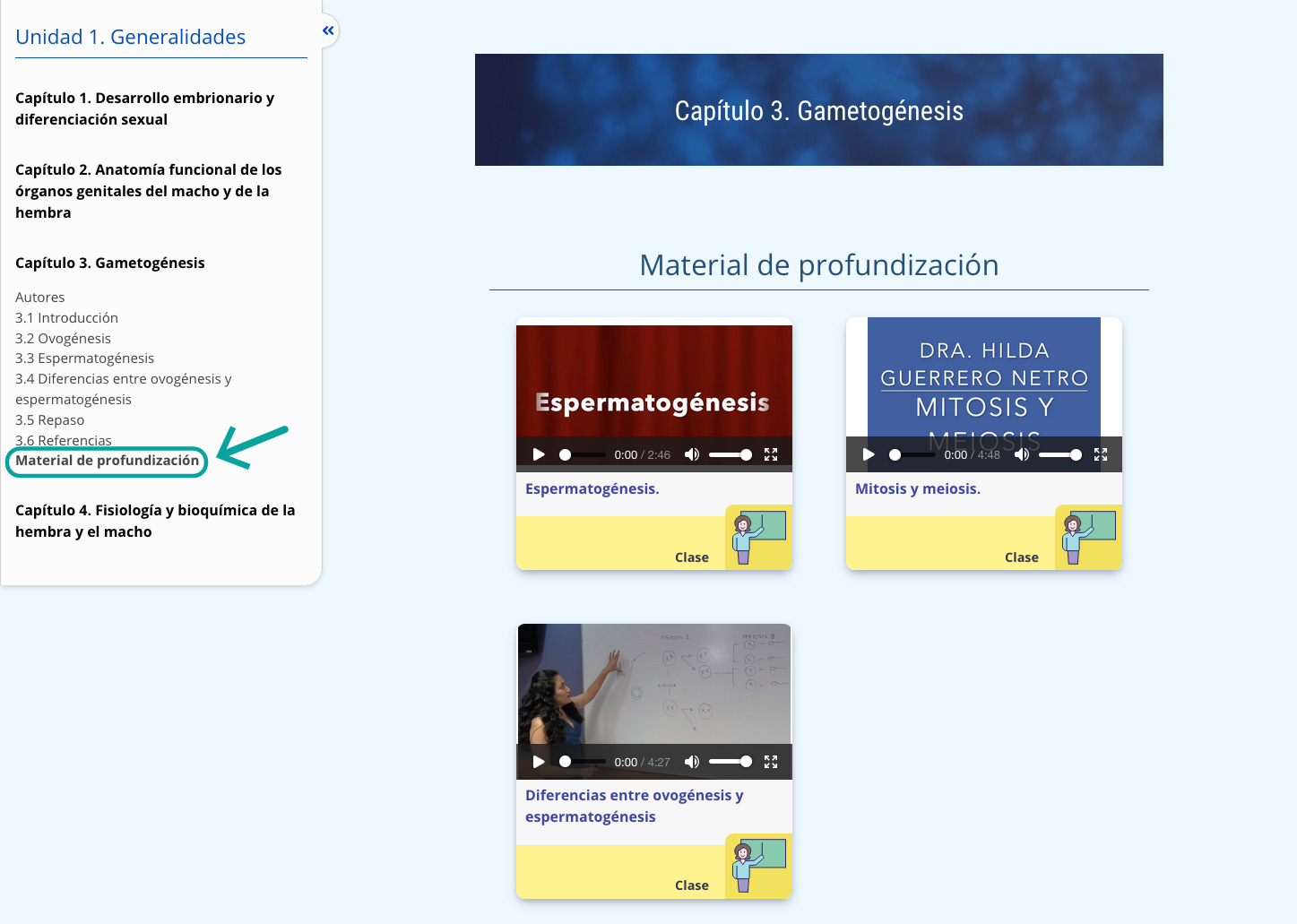
Para los espermatozoides depositados en el aparato genital de la hembra, la ZP es la última barrera que deben atravesar para fertilizar al óvulo. En la penetración de la ZP interviene un mecanismo mecánico y otro enzimático. En el primero las fuerzas generadas por el movimiento hiperactivado ayudan al paso del espermatozoide a través de la ZP. En el segundo, las enzimas liberadas durante la RA crean una abertura en la ZP, a través de la cual el espermatozoide puede penetrar. En la porción inmediata de la cabeza espermática, la superficie de la ZP parece erosionar, formando un orificio o camino espermático (figura 11).
La acrosina (ACR), liberada del espermatozoide durante la reacción acrosomal, tiene propiedades únicas. Ésta hidroliza las glucoproteínas de la ZP sin alterar el resto de la estructura de la ZP, ni los sitios de unión del espermatozoide a la misma. La acrosina se encuentra primero en su forma inactiva, la proacrosina. Ésta tiene una fuerte habilidad para unirse a carbohidratos, por lo tanto parte de la proacrosina liberada del espermatozoide durante la reacción acrosomal se une a la superficie externa de la ZP (específicamente a la glicoproteína ZP2 de la ZP) y otra parte permanece unida a la membrana acrosomal interna. La proacrosina en ambos sitios ayuda a mantener al espermatozoide con RA sobre la ZP, fenómeno al que se le da el término de “unión espermática secundaria”. Enseguida, la proacrosina por un mecanismo autocatalítico se convierte en su forma activa, la acrosina alfa, que tiene actividad proteolítica. La acrosina alfa "suaviza" la ZP, permitiendo que el espermatozoide con RA inserte su cabeza dentro de la ZP. Sin embargo, recientemente se ha mostrado que la proacrosina no es esencial para la penetración de la ZP, ya que en su ausencia, en el caso del ratón, los espermatozoides tienen la capacidad de transitar por la ZP, más aún se postula que su papel es dispersar la matriz acrosomal, por lo tanto otras enzimas acrosomales deben estar participando.
La penetración de la zona pelúcida se realiza en 10 minutos de manera in vitro, sin embargo, se cree que in vivo el proceso es más rápido. La penetración ocurre en trayectoria oblicua. Una vez atravesada la ZP, el espermatozoide fertilizante atraviesa rápidamente el espacio perivitelino. La cabeza espermática se une a la membrana plasmática del óvulo y rápidamente la cabeza es incorporada dentro del citoplasma del óvulo (video 2).
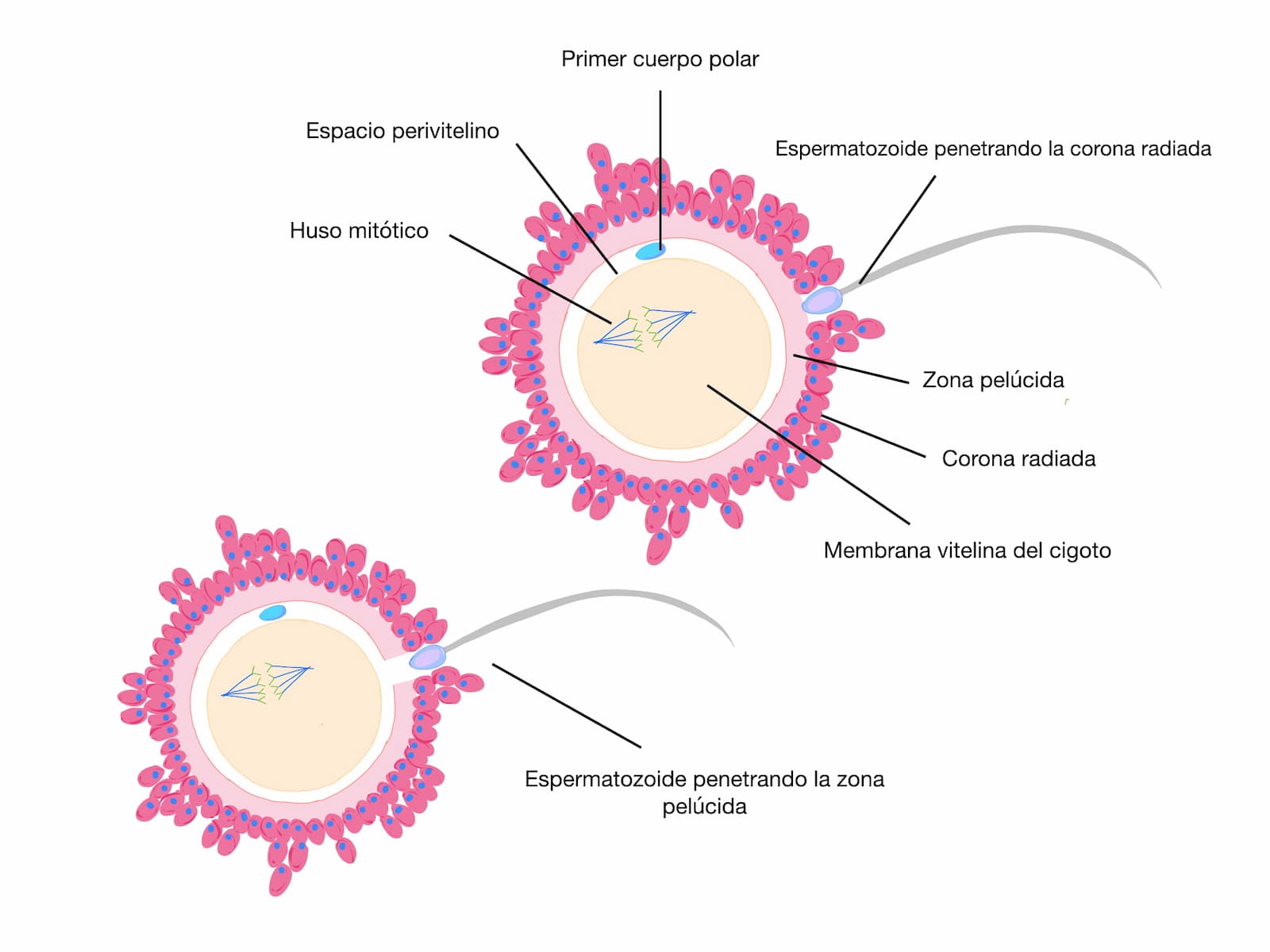
En el óvulo, un subdominio específico de la membrana plasmática es el que participa en la mezcla inicial de las dos bicapas lipídicas. La membrana plasmática del óvulo puede ser dividida en dos regiones principales, la parte de la membrana que cubre directamente los cromosomas metafásicos y que tiene una superficie lisa desprovista de microvellosidades; diversos investigadores han descrito que ningún espermatozoide se fusiona en esta región; el resto del óvulo es rico en microvellosidades, siendo esta región en donde el espermatozoide se une y fusiona.
Tanto en el óvulo como en el espermatozoide, la región de la membrana plasmática que participa en la fusión inicial está restringida. Las regiones excluidas de la fusión inicial son la membrana del flagelo y la recién incorporada membrana acrosomal interna. De manera consistente se ha postulado que solo la región ecuatorial del espermatozoide puede iniciar la fusión con el óvulo (figura 12), pero no se descarta que la membrana plasmática de la región posterior de la cabeza también lo haga. Así, varias proteínas espermáticas han sido propuestas en la mediación de la fusión del gameto masculino, entre ellas se encuentran: IZUMO1, cirestestina (que pertenece a una familia de proteínas de adhesión a matriz extracelular), CRISP1 (familia de proteínas secretada ricas en cisteína), CRISP2, SLLP (proteína espermática semejante a lisozima c), la ecuatorina y la familia de glicoproteínas ADAMs, entre ellas la Fertilina.
Conforme el espermatozoide es incorporado en el citoplasma del óvulo y las membranas de los dos gametos se mezclan para formar una membrana nueva en el cigoto, la membrana acrosomal interna es excluida de esta mezcla. Subsecuentemente, la región posterior de la cabeza y el flagelo son incorporados por el óvulo vía fusión de membranas, mientras que la región anterior de la cabeza es introducida mediante un sistema fagocítico. En el sitio de fusión de ambos gametos, el citoplasma forma una protrusión a menudo referida como “cono de fertilización”. Un indicador visible de la fusión espermatozoide-óvulo es la reducción súbita del movimiento flagelar. Bajo estas condiciones fisiológicas, sólo un espermatozoide se fusiona con la membrana plasmática del óvulo.
7.4.2 ACTIVACIÓN DEL OVOCITO Y FORMACIÓN DE PRONÚCLEOS
La respuesta del óvulo a la fusión de membranas entre ambos gametos es su activación. La activación completa del óvulo incluye la inducción temprana de cambios oscilatorios en la concentración del calcio intracelular y la reanudación de la meiosis, produciéndose un embrión capaz de desarrollarse normalmente.
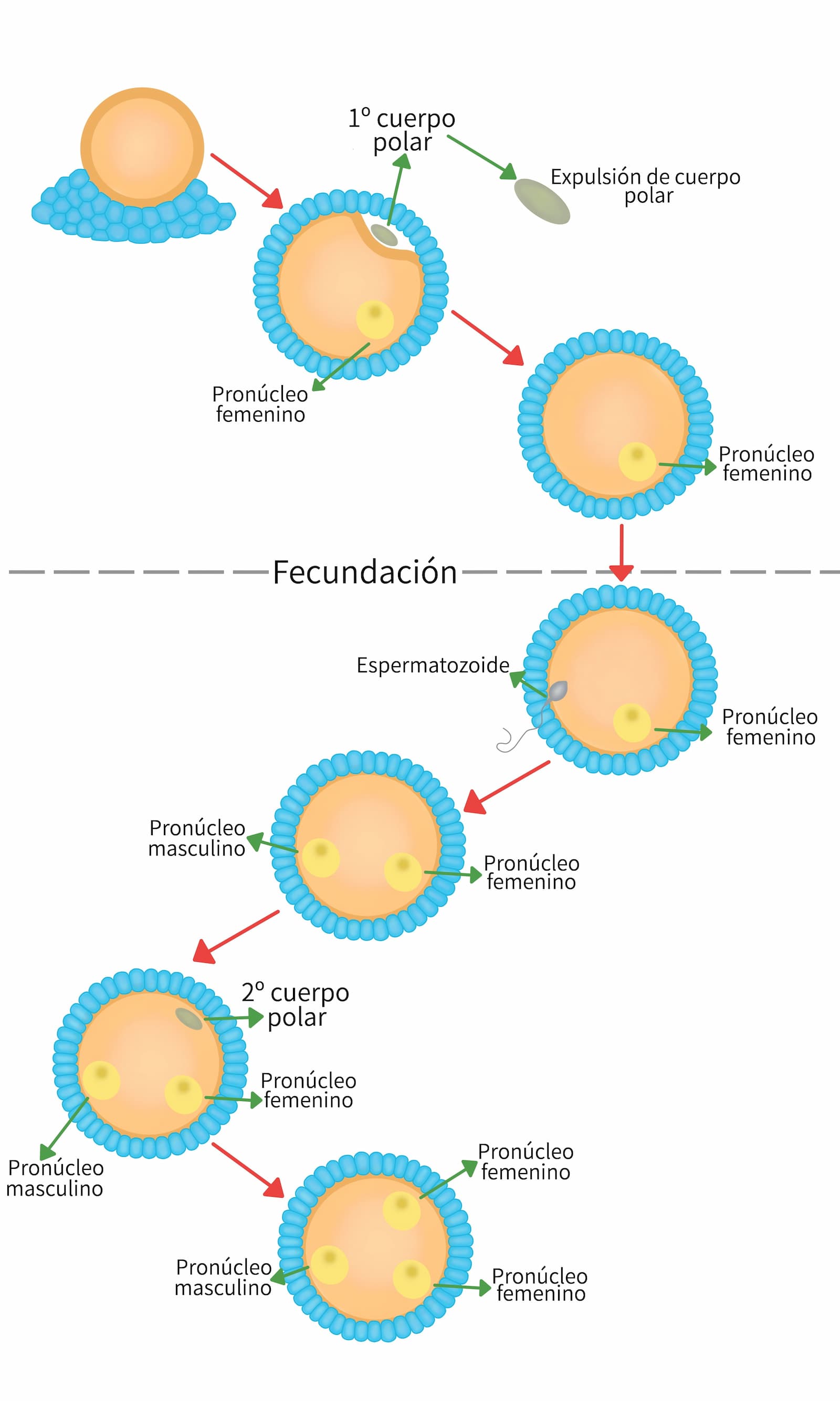
El óvulo en fase latente, en la metafase de la segunda meiosis termina su división, de tal manera que la mitad del contenido de cromatina es eliminado con el segundo cuerpo polar, evento que convierte al óvulo en una célula haploide.
La fusión de gametos precede la iniciación de las oscilaciones de Ca2+ en el óvulo, gracias a que el espermatozoide de mamífero contiene un factor soluble que se difunde dentro del óvulo enseguida de la fusión de los gametos, lo que luego inicia la liberación de Ca2+. Para ello, varias proteínas han sido involucradas: (1) la proteína de 33 kDa (oscilogEno), (2) la forma truncada del receptor c-kit, tr-kit, (3) la proteína de adhesión dominio WW de la hoja postacrosomal de la teca perinuclear espermática (PAWP) y (4) una fosfolipasa C específica del espermatozoide, llamada PLCzeta (PLCζ). Se piensa que probablemente esta última sea el factor espermático activador del ovocito; de manera singular en muchas especies de mamíferos se ha mostrado que PLCζ estimula las oscilaciones citoplásmicas de Ca2+ correspondiendo con aquellas de la fertilización y que disparan los eventos tempranos del desarrollo embrionario, ello en ratón, humano, rata, cerda, vaca e inclusive en otras especies no mamíferas.
El primer cambio visible que ocurre en el núcleo espermático, después de su incorporación en el citoplasma del cigoto, es el rompimiento de la envoltura nuclear estando la cromatina expuesta al citoplasma del cigoto. Eventualmente, la cromatina comienza a descondensarse hasta volverse una masa homogénea dispersa. Durante este proceso las protaminas son removidas del ácido desoxirribonucleico (DNA) y se reemplazan por histonas. Estos cambios son seguidos por el ensamblaje de una envoltura nuclear nueva alrededor de la cromatina descondensada para formar el pronúcleo masculino y se inicia la replicación del DNA.
Después de la anafase II, los cromosomas del cigoto permanecen dispersos, organizándose alrededor de estos una envoltura nuclear nueva, formándose el pronúcleo femenino y la replicación del DNA. Cada pronúcleo contiene un número variable de nucleolos.
7.4.3 SINGAMIA
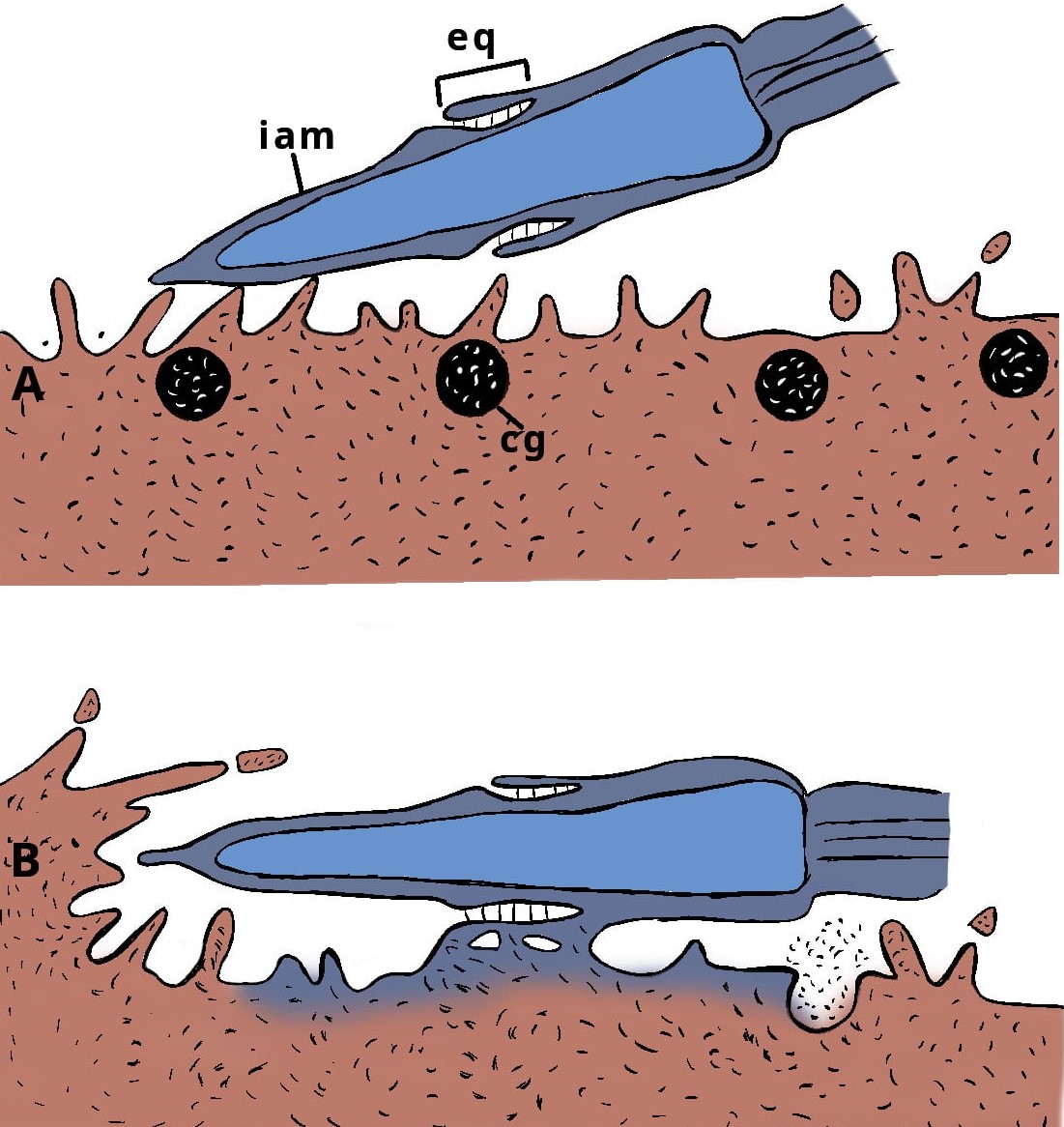
La secuencia de eventos que se dan durante la migración del pronúcleo femenino y del pronúcleo masculino hacia el centro del cigoto y su unión es lo que se conoce como singamia y se considera el punto de terminación de la fertilización y el inicio del desarrollo embrionario.
La síntesis de DNA inicia casi sincrónicamente en ambos pronúcleos. Para asegurar el desarrollo sincrónico de los pronúcleos, el cigoto posee una actividad supresora que previene el desarrollo del pronúcleo femenino durante las primeras horas de la activación del cigoto. Simultáneamente, el desarrollo del pronúcleo masculino parece también estar sincronizado con la terminación de la meiosis y el desarrollo del pronúcleo femenino. Conforme crecen los pronúcleos, se van acercando más uno al otro hasta que entran en contacto y se fusionan precisamente en el centro del cigoto, más tarde la totalidad de la envoltura nuclear se rompe dejando escapar una masa de cromosomas parcialmente condensados. Los cromosomas completan su condensación y se acoplan, uniéndose así las contribuciones hereditarias paterna y materna (interactivo 3). Esto es la profase de la primera división celular para dar origen a los dos primeros blastómeros.
Aposición de los pronúcleos.
Interdigitación de las superficies proximales de los pronúcleos.
Interdigitación de las superficies proximales de los pronúcleos.
Complejo de integración de los pronúcleos.
Vesiculación de las envolturas pronucleares y formación del huso mitótico.
Aparato mitótico de la primera metafase.
Los siguientes eventos pueden ser empleados como criterios de una fertilización exitosa (interactivo 4), aunque no todos son necesariamente apropiados para todas las especies:
- La reacción cortical (pérdida de los gránulos corticales del cigoto).
- Descondensación de la cabeza espermática dentro del cigoto.
- La presencia del primero y el segundo cuerpo polar.
- Formación y fusión del pronúcleo masculino y femenino.
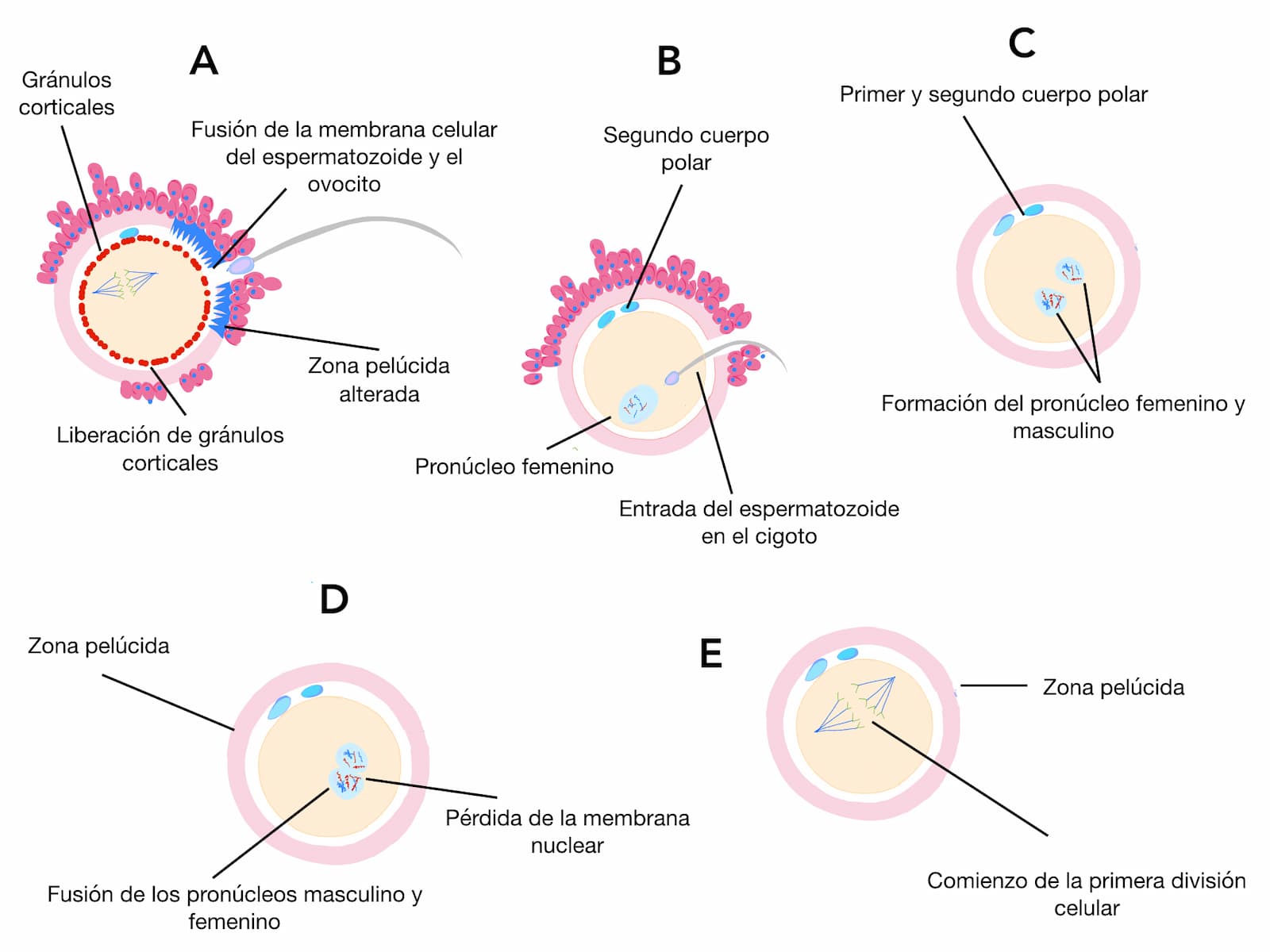
Penetración del espermatozoide a través de la zona pelúcida. Los gránulos corticales comienzan a disminuir.
Contacto del espermatozoide con la membrana plásmatica del cigoto. La cabeza del espermatozoide se hunde en el citoplasma. Se termina la 2a. división meiótica y se expulsa el 2o. cuerpo polar. La cola del espermatozoide tambien penetra. Se forma el pronúcleo femenino.
Los pronúcleos se desarrollan en forma sincronizada, el DNA se replica y empieza la transcripción de los genes maternos y paternos llegando a su punto máximo durante la segunda segmentación.
Unión de los pronúcleos masculino y femenino.
Inicia la primera división embrionaria.
7.4.4 ERRORES DE FERTILIZACIÓN
-
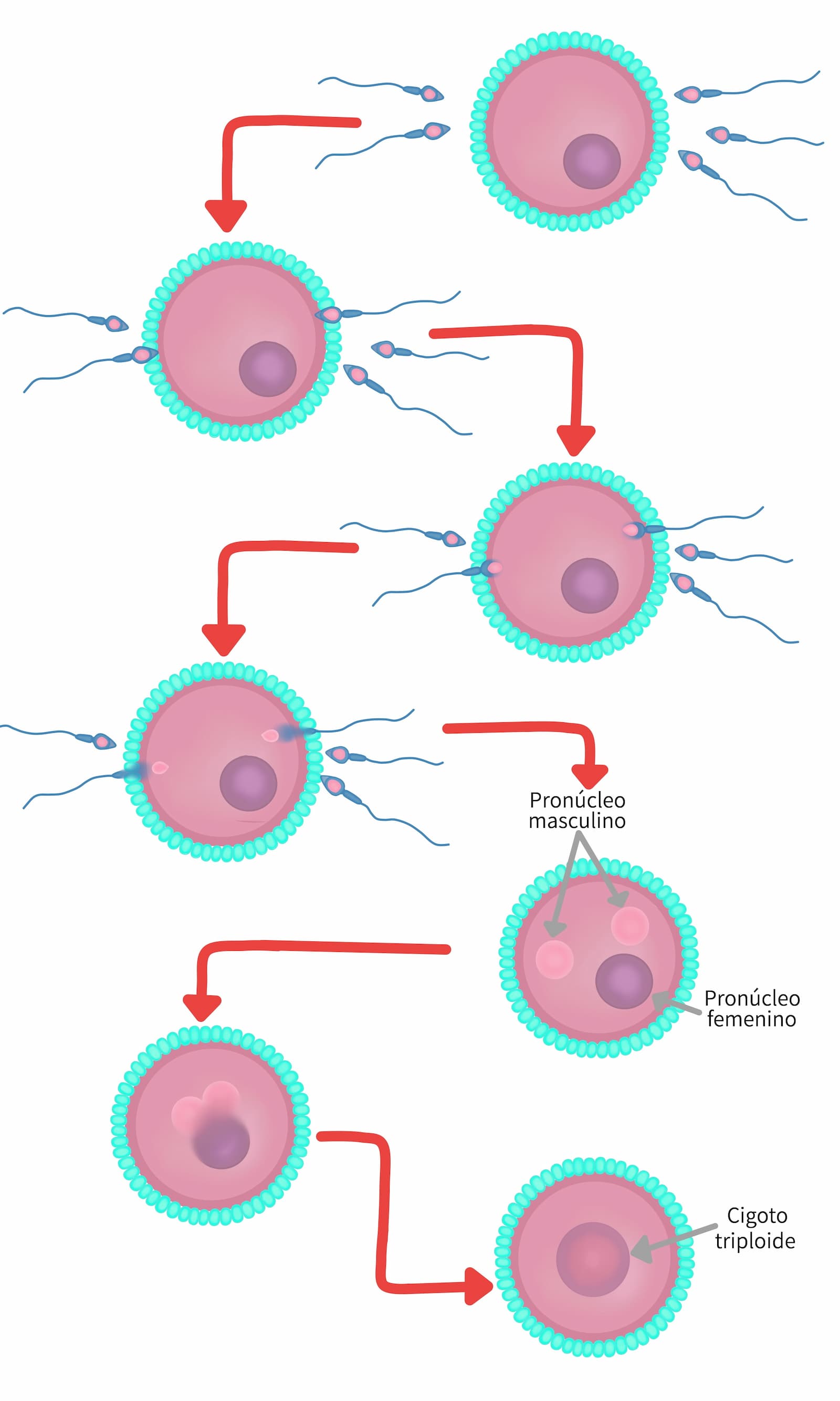
Poliespermia. Ocurre cuando 2 (excepcionalmente más) espermatozoides penetran al óvulo y ambos toman parte en la fertilización. Se forman tres pronúcleos (que no crecen tanto como en los casos normales) y durante la singamia los tres se fusionan. El cigoto tiene entonces 3 juegos de cromosomas y se llama triploide (figura 13). El desarrollo inicial del embrión es normal, pero posteriormente, degenera y muere. Por lo mismo la poliespermia es una condición letal.
Este problema puede incrementarse por cambios asociados al envejecimiento del óvulo antes de ser fertilizado, de ahí la importancia de que la monta o la inseminación artificial se realice en el momento propicio. En la cerda, la monta tardía aumenta la incidencia de poliespermia.
Como respuesta del óvulo a la fecundación, para prevenir los efectos letales de la poliespermia se activan mecanismos específicos de bloqueo, estos consiguen mediante la pérdida de permeabilidad de la ZP y alteraciones en la superficie de la membrana, que se impida la penetración de más espermatozoides. Aunque el mecanismo exacto se desconoce, uno de los primeros eventos que ocurren al momento de la fecundación es una hiperpolarización transitoria de la membrana plasmática del óvulo. Este bloqueo de la membrana plasmática se establece un minuto después de la unión espermatozoide-óvulo. Poco después, los gránulos corticales son exocitados liberando su contenido (enzimas hidrolíticas y proteasas) hacia el espacio perivitelino y éste es responsable de modificar la ZP3 y de hidrolizar parcialmente a la ZP2 (reacción zonal). La exocitosis de los gránulos corticales (reacción cortical) inicia dos minutos después de la unión espermatozoide-óvulo. Ya que la membrana de los gránulos exocitados se integra a la membrana plasmática del cigoto, se especula que tanto su membrana como su contenido contribuyen al establecimiento del bloqueo a la poliespermia. Cuando estos mecanismos fallan, entonces se tienen los casos de poliespermia.
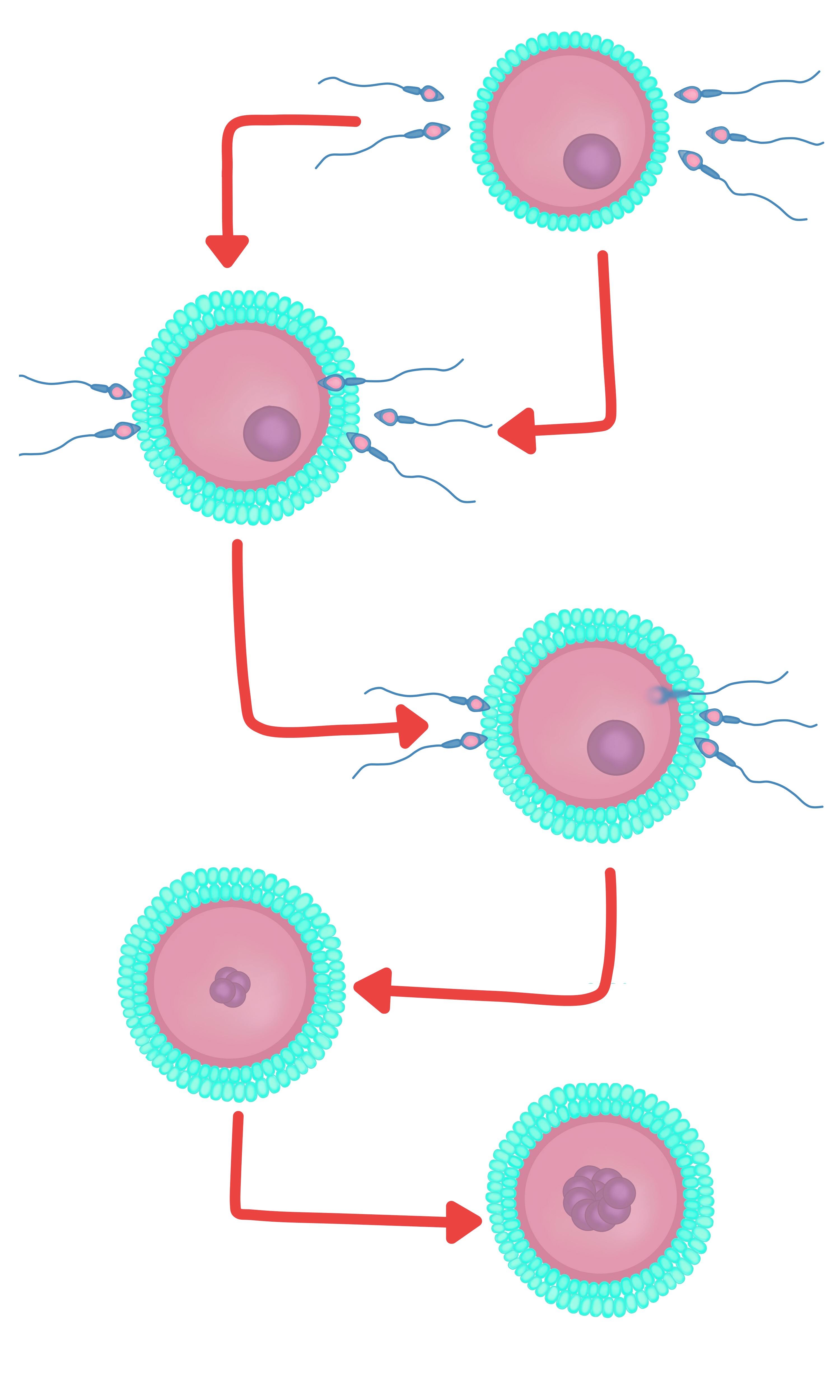
- Poliginea. Ocurre cuando el segundo cuerpo polar no es eliminado y se desarrollan entonces dos pronúcleos femeninos y uno masculino. Se forma igualmente un cigoto triploide y muere alrededor de la mitad de la gestación (figura 14).
- Ginogénesis. Este fenómeno se presenta en algunos peces (Molly amazónico). En ellos no hay machos, sólo hembras. Las hembras copulan con machos de una especie bisexual del mismo género. Los espermatozoides al penetrar el óvulo, sólo activan el cigoto ya que degeneran y no llegan a formar el pronúcleo masculino. El óvulo no expulsa el segundo cuerpo polar, y sigue su desarrollo como un ente haploide (figura 15). Esto no ocurre en mamíferos.
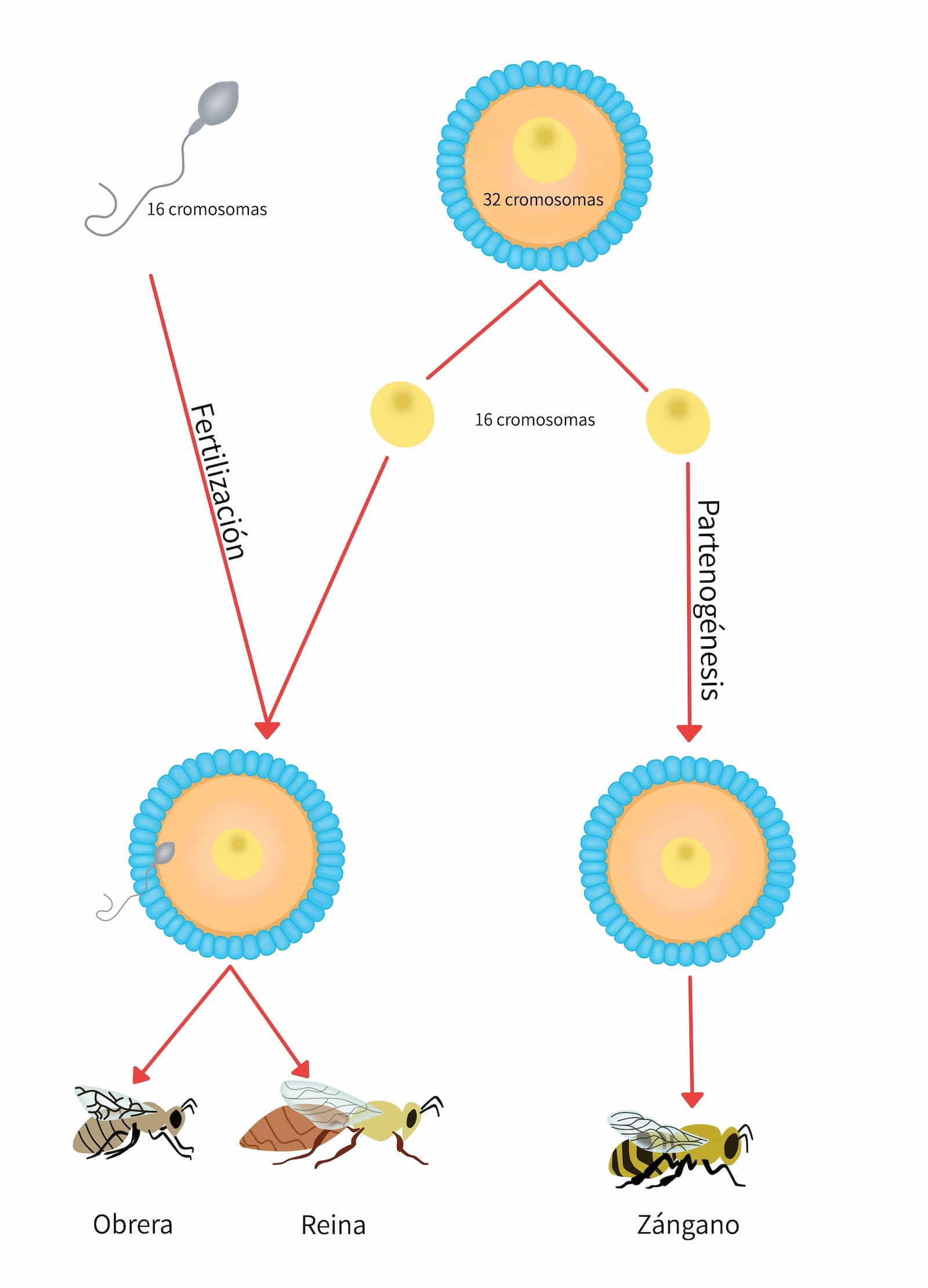
- Partenogénesis. Significa “parto de una virgen” y se aplica el término al desarrollo embriológico sin la participación de espermatozoide. Debe diferenciarse de ginogénesis donde el espermatozoide es necesario para activar el proceso. Este es un proceso normal en algunos insectos como la abeja, en que los zánganos son producidos por partenogénesis y las obreras y la reina, por fertilización (figura 16). En mamíferos, la partenogénesis avanza solamente hasta el estadio de implantación. Sin embargo, se piensa que dentro de la población humana podrían existir algunos productos de la partenogénesis. Naturalmente tales productos serían invariablemente hembras y se parecerían mucho a la madre. La partenogénesis ocurre con frecuencia en pavos y gallinas dando lugar a machos diploides.